只有尾巴的"噬菌体"还能杀菌吗?
对噬菌体有所了解的朋友们应该都清楚,有尾噬菌体的结构主要包括由包裹着遗传物质的衣壳蛋白组成的头部和长尾、肌尾、短尾等不同形态的尾部结构。那如果没有了头部、只保留尾部的“噬菌体”还会有杀菌的效果吗?它们和正常结构的噬菌体之间又有什么样的进化关系呢?
1、什么是噬菌体尾部样细菌素(PTLBs)?
简单点说,可以认为PTLB是一种缺陷型的溶原噬菌体,它们是噬菌体尾部的结构同源物。PTLB广泛存在于细菌基因组中,随着时间的推移,它们已经独立进化并可以产生细菌素。PTLB是由丝裂霉素C通过细菌的DNA损伤或SOS反应诱导的,它通过释放diffocin和破坏膜电位杀死竞争细菌(图1)。根据其结构特点,PTLB可以被分为R型和F型,R型的结构特征是一个非柔性的可收缩纳米管,与肌尾噬菌体的尾部结构类似;F型(柔性型)具有柔性非收缩杆状结构,与长尾噬菌体的尾部结构类似。
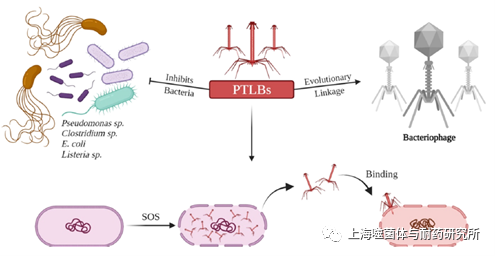
图1. PTLB和噬菌体
2、PTLB与噬菌体的进化关系
不同的噬菌体尾部结构与不同类型的PTLB相似,比如噬菌体PS17.TP901-1和 lambda的尾部分别类似于R型pyocins、F型monocins和F型pyocins(图2)。结构上的相似性说明PTLB和噬菌体尾部之间存在着进化关系。例如丁香假单胞菌噬菌体PS17和R型pyocin有相似的结构,都有6根尾丝蛋白附着在尾管结构上;R型pyocin识别的受体是与可以与PS17发生相互作用的脂多糖;这两者在形态结构和受体特异性表明它们具有共同的特征。Nakayama等人通过分析位于PAO1染色体上核苷酸序列研究了噬菌体和pyocin之间的关系,结果显示R型pyocin与P2噬菌体有对应关系,F型pyocin与lambda噬菌体有对应关系。通过诱导SOS反应可产生和释放PTLB,这一点也和溶原噬菌体类似,同样支持了它们来自缺乏衣壳基因和复制能力的缺陷型前噬菌体的事实。目前普遍认为人们PTLB在形成后发生了各种各样的适应性进化,使得它们具有更有效地破坏细胞膜电位的能力。
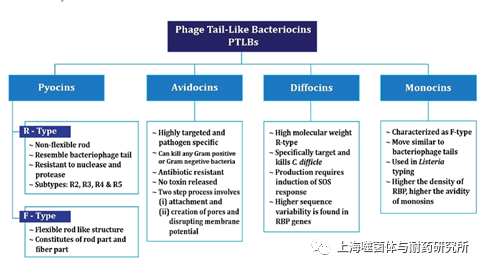
图2. PTLB的分类和特点
3、PTLB的作用机制
PTLB是一种导致宿主细胞膜去极化和杀死细胞的窄谱抗菌剂。PTLB和噬菌体共有的关键决定因素之一是受体结合域(RBP),识别细胞表面特定的受体,如蛋白质、多糖(脂多糖、荚膜多糖),菌毛或鞭毛等结构。R型PTLB的杀菌机制类似于肌尾噬菌体将DNA递送到细胞内的过程。R型PTLB的作用机制包括通过尾丝(tail fiber)中的RBP与细胞表面的受体结合,导致尾鞘收缩;随后,它通过尾针蛋白(tail spike)穿过细胞膜将内核强行穿过宿主细胞膜。形成通道导致的离子流动,破坏了细胞膜内的离子浓度梯度,最终导致细胞死亡(图3)。F型不具有收缩的功能,因此不能直接穿透细胞膜,它们通过分泌某种单一粒子在细胞膜上形成通道,从而扰乱细胞呼吸和膜电位来杀死细菌。
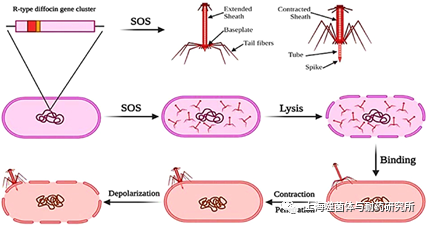
图3. PTLB的杀菌机制
小结
PTLB是具有一定应用潜力的抗菌物质,也是研究的热点方向。它们在细菌里面广泛存在并且方便进行基因工程改造。PTLB是不含噬菌体衣壳和遗传物质的,它们不是生命体,而是一种大分子蛋白类物质。但是它们的杀菌机制与噬菌体识别和吸附宿主的过程类似,都有很强的特异性,因此也会存在一个宿主谱窄的问题。目前有关PTLB的研究还都停留在实验室水平,没有临床试验结果的支持。所以,具体未来前景如何,我们可以期待一下!
参考文献
[1]https://pubmed.ncbi.nlm.nih.gov/36162371/
1、凡本网所有原始/编译文章及图片、图表的版权均属微生物安全与健康网所有,未经授权,禁止转载,如需转载,请联系取得授权后转载。
2、凡本网未注明"信息来源:(微生物安全与健康网)"的信息,均来源于网络,转载的目的在于传递更多的信息,仅供网友学习参考使用并不代表本网同意观点和对真实性负责,著作权及版权归原作者所有,转载无意侵犯版权,如有侵权,请速来函告知,我们将尽快处理。
3、转载请注明:文章转载自www.mbiosh.com
联系方式:020-87680942