luxS在空肠弯曲杆菌中的作用 超越细胞间信号传导

一、文章背景:
在空肠弯曲菌中,luxS 基因的转录与几种表型相关,包括细胞间信号传导、运动、生物膜形成、宿主定植、毒力、自凝集、细胞粘附和侵袭、氧化应激和趋化性(图1)。已经表明,luxS突变体中完全不存在细胞间信号传导,而与野生型相比,运动性,生物膜形成,自凝集,宿主定植和luxS突变体的侵袭减少。
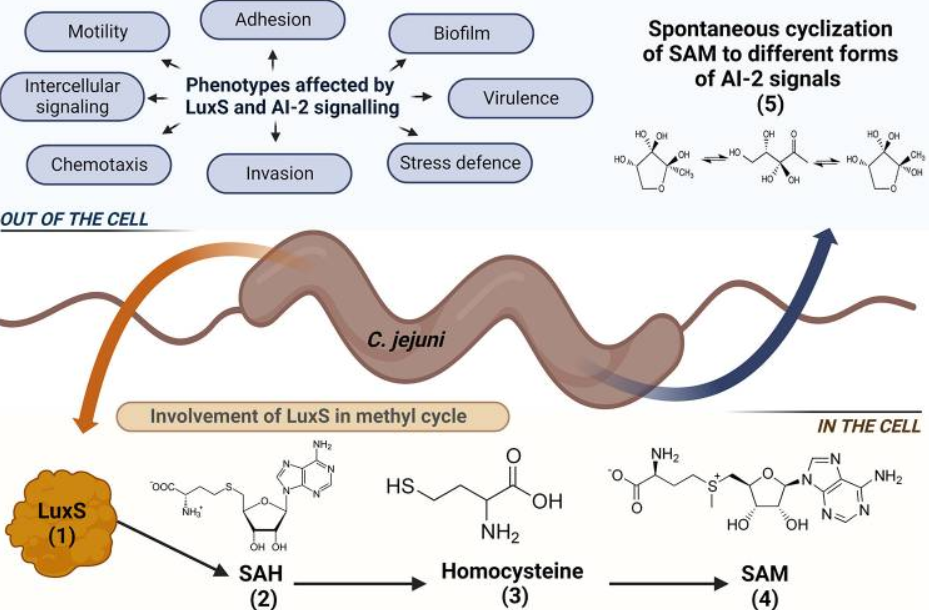
图1 LuxS蛋白在空肠弯曲菌中的作用示意图。LuxS在细菌细胞中的主要功能是S-腺苷-同型半胱氨酸循环途径,它是甲基循环的一部分,它负责S-腺苷同型半胱氨酸(SAH; 2)水解为同型半胱氨酸,然后进一步代谢为SAM。SAM 途径的产物之一也是 4,5-二多面体氧基-2,3-戊二酮 (DPD),它自发环化为 AI-2,一种种间信号分子。空肠衣原体中受luxS和AI-2信号传导影响的表型包括细胞间信号传导、运动性、粘附性、生物膜形成、趋化性、侵袭、毒力和应激防御。
尽管很明显luxS基因对空肠弯曲菌的许多表型很重要,但luxS在其他生物过程中的作用,例如不同必需氨基酸的合成以及氮和丙酮酸代谢,迄今为止尚未明确描述。因此,作者使用了不同水平的综合方法,包括生理学,转录组学和蛋白质组学,以研究luxS基因的关键作用,比较了空肠弯曲菌11168Δ luxS(luxS突变体)与空肠弯曲菌NCTC 11168(野生型)。
二、文章方法:首先,luxS突变型和野生型菌株用于评估应激条件下(饥饿和氧化应激)下的存活和形态差异。还研究了它们对不同碳源和氮源的利用。此外,作者进行了二代测序,以深入了解luxS突变株和野生型菌株之间的转录组学差异。此外,无标记相对定量质谱法用于所选菌株的蛋白质组学分析。本文首次尝试在分子水平上研究luxS基因参与空肠弯曲菌过程。
三、文章结果:
1.luxS突变型和野生型菌株在胁迫条件下的存活。
luxS突变体和野生型空肠弯曲菌在最佳条件(最佳气氛,培养基和温度)下培养,以确定其生长过程中可能出现的基本差异。此外,还研究了luxS突变型和野生型菌株在饥饿(低营养)和氧化应激(有氧气氛)条件下的存活。在整个栽培期间,两种菌株在最佳条件下的存活率差异均无统计学意义(P ≥ 0.05)(图2)。在每个时间点,野生型对饥饿(P ≤ 0.05)比luxS突变体更敏感(图2).这些差异超过3个log10CFU/mL。氧化应激对两种菌株的存活均无显著影响,但120 h后,野生型的存活率为1 log10CFU/mL 高于 luxS 突变体 (P ≤ 0.05)(图2)。

图2(甲至丙)空肠弯曲菌NCTC 11168和空肠弯曲菌11168Δ luxS(luxS突变体)在最佳(A)、饥饿(B)和氧化(C)条件下在42°C条件下孵育120 h期间的生长速率。数据是平均Log10CFU值,并表示为三个重复±标准偏差的平均值;*,P ≤ 0.05 与相关对照组
3.用于透射电子显微镜(TEM)的生物样品的负染色制备提供了有关细菌形状和大小的信息,作者跟踪了饥饿和氧气的存在对空肠弯曲菌野生型和luxS突变株形态的影响。在正常生长条件下,空肠弯曲菌野生型和luxS突变体表现出两种细菌形态型的混合:螺旋(图3A) 和曲率形状的杆 (图3B)有两根鞭毛,每侧一根。虽然螺旋形状被描述为空肠弯曲菌的正常形状,但它们的形态可以转化为杆状。两种形状都存在于两种菌株中,但在luxS突变体中,杆状占上风,并且与野生型(1.96μm×0.67μm)相比,细胞较小(2.77μm×0.61μm)。饥饿和氧化应激在两种菌株中引发了相同的反应。螺旋形和曲率形状转化为球形(图3C)在两种胁迫条件下,以及暴露5天后,作者只能找到球状体。在暴露于氧应激后立即(4小时),作者在luxS突变体的样品中找不到任何保存的细胞;仅存在球状形态。
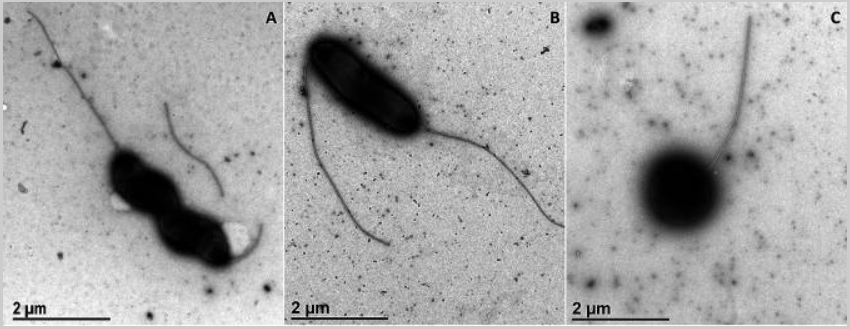
图3 空肠弯曲菌形态学的代表性显微照片。(A) 螺旋形,空弯NCTC 11168。(B)杆形,空弯11168Δ luxS(luxS突变体)。(C)球形,空肠弯曲菌11168Δ luxS(luxS突变体);比例尺,1 μm。
4.luxS突变型和野生型对不同碳和氮源的利用。
使用Biolog系统获得的结果表明,与野生型相比,luxS突变体仅对22种不同碳和氮源中的288种代谢不良(P ≤ 0.05)(图4).当暴露于l-苹果酸,l-脯氨酸,l-丝氨酸,富马酸,柠檬酸,l-谷氨酰胺,l-天冬酰胺,l-岩藻糖等时,从野生型收集的信号比luxS突变体高10倍,这意味着luxS突变体的代谢在暴露于这些营养物质后受到限制(P ≤0.05)。此外,这意味着luxS突变体在这些来源上的生长速度和存活能力低于野生型,野生型在这22个来源上的生长优于luxS突变体。

用Biolog系统测量的不同碳和氮源的空肠弯曲菌NCTC 11168和空肠弯曲菌11168Δ luxS(luxS突变体)利用之间的差异。数据表示为与luxS突变体相比,从野生型收集的信号的倍数变化。
luxS突变型和野生型的转录组学图谱。
5.比较野生型和luxS突变型空肠弯曲菌的转录组可以提高作者对luxS基因在空肠弯曲菌生命周期中的作用的理解。菌株一直培养到指数阶段的中间,在此期间预计基因表达会发生许多变化。详细分析表明,孵育765 h后,归一化为野生型(假发现率[FDR],P ≤ 0.05)时,luxS突变体中差异表达16个基因。在这些基因中,354个上调,402个下调(无花果。5;表 S1)。然而,选择这些转录本的列表(表S1)是因为它们在luxS突变体和野生型之间显着不同(log2倍数变化 [FC] 为 ≥1,P ≤ 0.05)。下面列出了其中一些基因,因为它们可以在生理水平上解释作者的观察结果,例如在类似饥饿的条件下生长。在上调基因中,检测到对空肠弯曲菌合成代谢重要的基因,包括lys-C,aspS,pyrB,glnP,glnS,proA,gltB,argC,hemA和ilvI。在下调基因中,检测到对空肠衣原体分解代谢重要的基因,包括htrA、pyk、Cj0021c、Cj1418c、Cj1417c、purQ、aspB、Cj0073c和fcl (表1).luxS突变体中差异表达基因的网络表明,大多数基因包含在生物过程中,如三羧酸(TCA)循环,丙酮酸,氮和硫胺素的代谢以及脂多糖的生物合成,并且这些生物过程上调。许多其他途径,包括鞭毛组装、ABC转运蛋白和蛋白质转运等双组分系统,也被上调,而许多核糖体基因被下调(无花果。6).在参与鞭毛运动和非生物表面定植的30个差异表达基因中,有22个上调,包括flhF,flgI,flgB,flaG,flgM,flgD,flgG2,flgE2,fliD,flgK,fliS,flaA,pseB,pseC,ptmA,maf4,legF,Cj1319, cj1330、cj1026c、cj0391c 和 cj0977。 此外,许多对应激反应很重要的基因要么上调(cstA,tpx,trxA,trxB,kata,fdxA和ahpC),要么被下调(grpE,dnaK和htpG)。
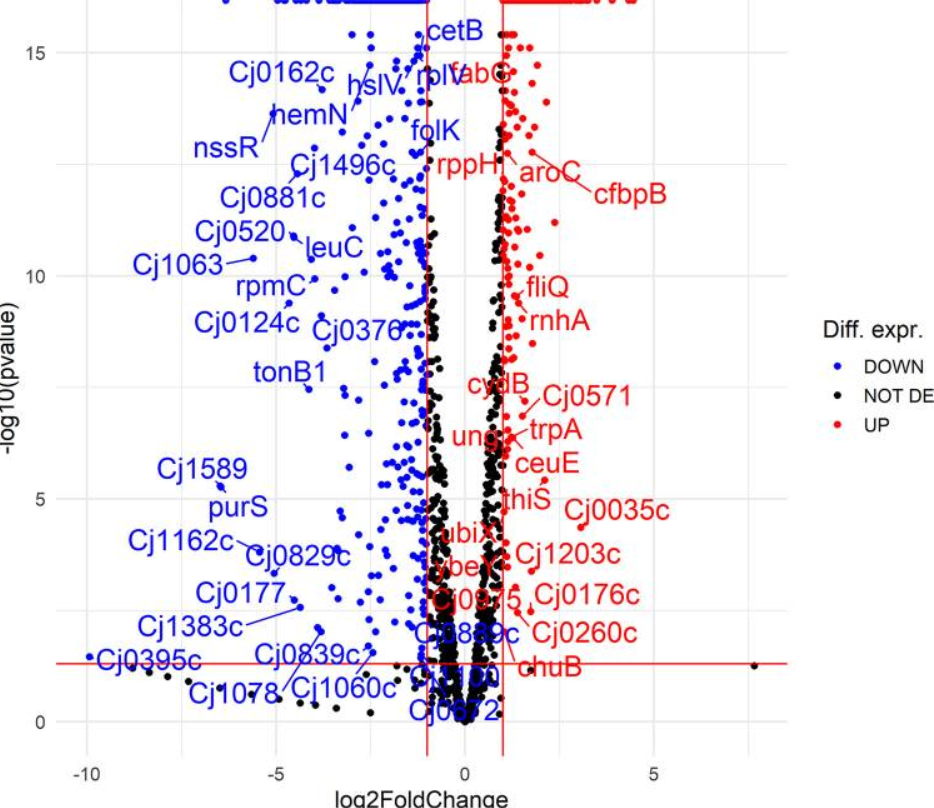
火山图显示空肠弯曲菌NCTC 11168和空肠弯曲菌11168Δ luxS(luxS突变体)的转录组学数据。
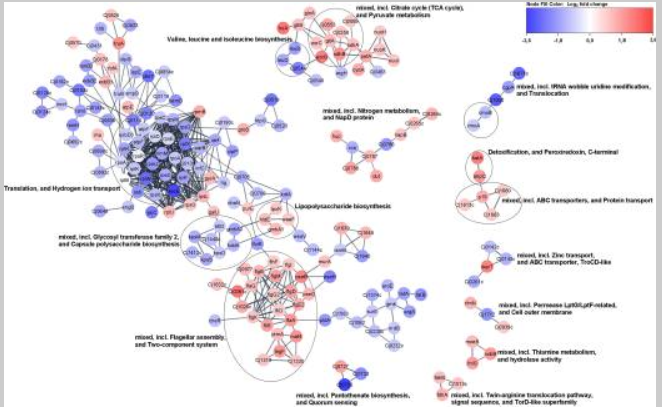
空肠弯曲菌11168Δ luxS(luxS突变体)中差异表达基因的网络。
此外,与野生型相比,对luxS突变体中显着差异表达的基因(FC ≥ 1,P ≤ 0.05)进行了基因本体(GO)富集分析,并显示出以下结果(图7):
(一)生物过程,包括氧化还原过程,电子传递链和呼吸电子转运链,差异表达,但没有具体说明过程是上调还是下调。(二)分子功能上调二级活性跨膜转运蛋白活性、共转运蛋白活性、溶质:阳离子共转运蛋白活性、溶质:钠共转运蛋白活性、转运蛋白活性、离子结合、辅因子结合、氧化还原酶活性(作用于NAD[P]H)和氧化还原酶活性(作用于NAD[P]H、醌或类似化合物),并下调rRNA结合。
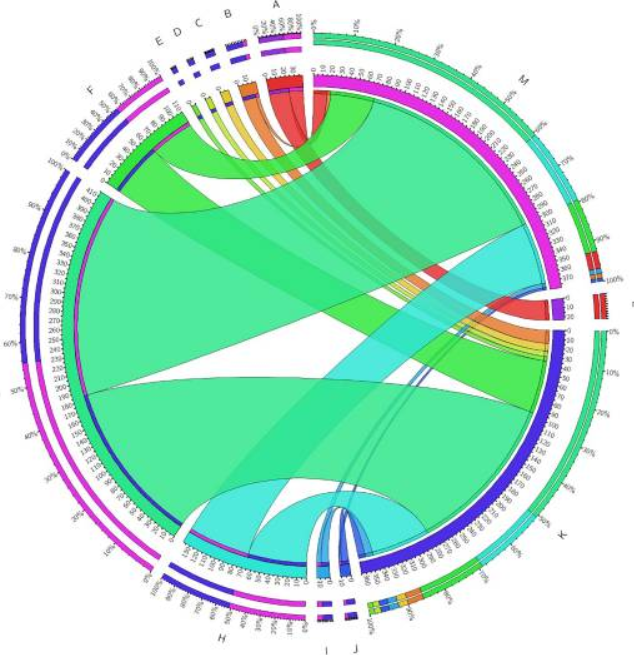
空肠弯曲菌11168Δ luxS(luxS突变体)分子功能的功能分类
8.luxS突变型和野生型的蛋白质组学特征。
进一步分析了蛋白质组学谱,通过比较野生型和luxS突变菌株来确认luxS在空肠衣原体生命周期中的作用。详细分析显示,样品中总共鉴定和定量了206种蛋白质。在这些蛋白质中,有53种差异表达(FDR,P≤0.05)(图8).正如预期的那样,最大的倍数变化属于luxS,反映了突变株中缺失突变的有效性。与其他52种差异表达蛋白中,与野生型相比,32种在luxS突变体中上调,20种下调(FDR,P ≤ 0.05)(图8).

火山图显示空肠弯曲菌 11168ΔluxS(luxS 突变体)与野生型相比的蛋白质组学数据
具体而言,与野生型相比,luxS突变体中所选蛋白质在统计学上存在差异(P ≤ 0.05)。luxS突变体中的上调蛋白由在合成代谢中起重要作用的组(FolE,DapB,Dcd,HemE,ThiG,AroB,PurA,IlvC,GuaA,PurD,DapE,LeuS,HisA,FabH和PseI),分解代谢(GpmI和Eno),细胞分裂(TolB和ParB)和粘附(PorA)组成。下调蛋白在合成代谢(Prs,LegG,LegI,DapA,ThyX和AcpP),分解代谢(Cj1418c),DNA修复(RadA和TopA),细胞壁形成和组织(MurB和LpxB)以及蛋白质修饰(HypA,SelD和PepA)中起重要作用。与野生型相比,对luxS突变体中显着差异表达的蛋白质进行了GO富集分析,并显示出以下分子功能差异(图9).上调分子功能主要包括与另一个分子上的一个或多个特定位点(ATP结合,核苷酸结合和金属离子结合)和催化活性基团的选择性分子相互作用。最重要的下调分子功能是金属离子结合基团、转移酶活性基团和DNA结合基团。
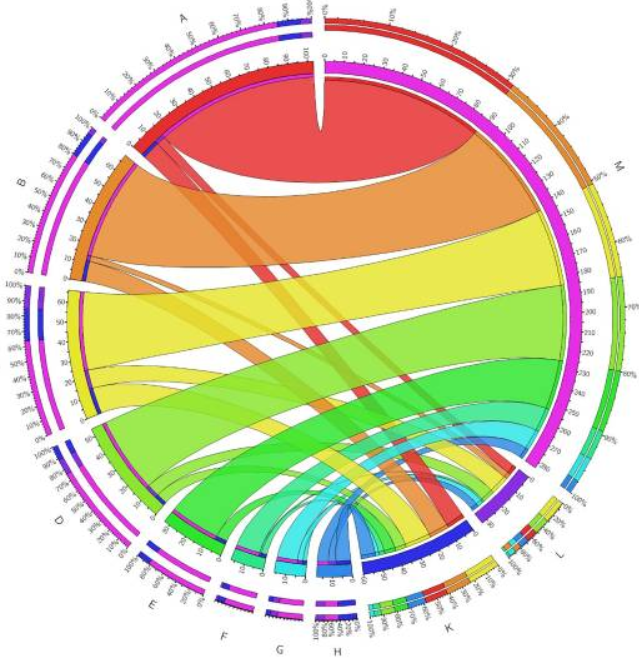
空肠弯曲菌11168Δ luxS(luxS突变体)中分子功能的功能分类。
差异表达蛋白(图10) 表示哪些生物学功能在 luxS 突变体中受到影响。上调蛋白被归入具有生物学功能的注释簇,包括核苷酸、氨基酸和次级代谢物的生物合成、单碳代谢、新蛋白的翻译、应激诱导的多伴侣系统和细胞粘附。下调的生物学功能包括DNA修复机制,一般应激反应,细胞壁组织以及生物合成和蛋白质修饰过程。

空肠弯曲菌 11168ΔluxS(luxS 突变体)中差异表达的蛋白质网络。
四、文章总结
转录组学和蛋白质组学分析揭示了 TCA 循环、丙酮酸、氮和硫胺素的代谢以及 luxS 突变型和野生型之间的脂多糖生物合成的主要生物学差异。这些过程被上调,可能是为了luxS突变体的自我保护。这些新发现表明,luxS突变体比野生型对压力条件做好了更好的准备。luxS突变体在饥饿条件下存活得更好,并且几乎与野生型一样利用了各种碳和氮源。此外,这里介绍的结果显着补充了最近非常重要的发现,即AI-2信号分子(luxS基因的产物)的浓度随着细胞浓度的增加而线性增加。因此,作者证实了luxS在中枢代谢途径中的强烈参与,超出了真正的细胞间信号系统。
参考文献:Ramić D, Jug B, Šimunović K, Tušek Žnidarič M, Kunej U, Toplak N, Kovač M, Fournier M, Jamnik P, Smole Možina S, Klančnik A. The Role of luxS in Campylobacter jejuni Beyond Intercellular Signaling. Microbiol Spectr. 2023 Feb 1;11(2):e0257222. doi: 10.1128/spectrum.02572-22. Epub ahead of print. PMID: 36722966; PMCID: PMC10100756.
 上一篇:六种食源性致病菌,致病菌的危害
上一篇:六种食源性致病菌,致病菌的危害- 下一篇:蜡样芽孢杆菌-重要的食源性致病菌
1、凡本网所有原始/编译文章及图片、图表的版权均属微生物安全与健康网所有,未经授权,禁止转载,如需转载,请联系取得授权后转载。
2、凡本网未注明"信息来源:(微生物安全与健康网)"的信息,均来源于网络,转载的目的在于传递更多的信息,仅供网友学习参考使用并不代表本网同意观点和对真实性负责,著作权及版权归原作者所有,转载无意侵犯版权,如有侵权,请速来函告知,我们将尽快处理。
3、转载请注明:文章转载自www.mbiosh.com
联系方式:020-87680942

