
细菌逆转录酶合成多聚腺苷的长链cDNA用于抵御噬菌体


作者探讨了大肠杆菌中DRT9型反向转录酶抗噬菌体防御系统的功能机制。研究发现,当将DRT9基因及其上游非编码RNA(ncRNA)引入大肠杆菌时,可以赋予细胞强大的抗噬菌体能力,尤其对Tevenvirinae噬菌体具有10-106倍的杀伤效果。这种抗性主要通过诱导感染细胞进入生长停滞状态,但不会导致大规模细胞死亡。随后,这些生长停滞的细胞能迅速恢复正常生长,CFU数目可增加105倍。

进一步研究发现,DRT9蛋白与其上游133nt的ncRNA形成稳定复合物,这种寡聚体复合物才是DRT9发挥抗噬活性的关键形式。同时,ncRNA及DRT9蛋白中保守的YADD催化位点都是抗噬活性的必需组分。这些发现揭示了DRT9依赖于与特定ncRNA形成复合体,通过诱导感染细胞进入可逆的生长停滞状态,从而达到有效抑制噬菌体感染的独特机制。
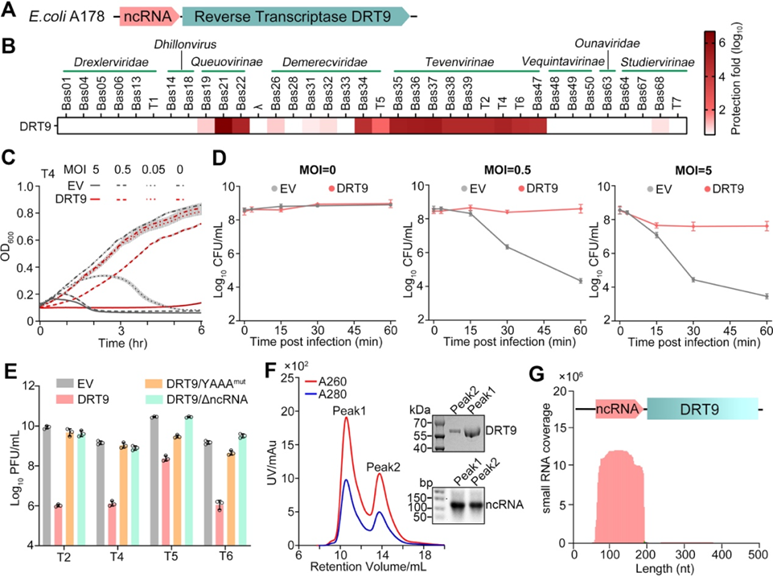
图1 DRT9与ncRNA结合,通过流产感染赋予大肠杆菌噬菌体防御能力
通过冷冻电子显微镜研究,作者发现DRT9-ncRNA形成稳定的六聚体复合物,其结构呈现"三聚二聚体"的配置。在这个六聚体复合物中,每个DRT9蛋白与一段约177nt的ncRNA紧密结合,形成一个封闭的构象。DRT9蛋白本身具有与其他逆转录酶相似的保守结构域,如手指结构域、掌状结构域和拇指结构域。但它还具有独特的C端尾巴,该尾巴与ncRNA相互作用,有助于稳定复合物。这段177nt的ncRNA呈"树状"结构,包括6个茎环结构(SL1-SL6)。其中,SL1和SL6形成树干,SL2-SL5则缠绕在DRT9的拇指结构域和C端尾巴周围,形成树枝状构象。这种DRT9与长ncRNA紧密结合,形成封闭构象,与此前报道的逆转录酶明显不同,可能是DRT9特有的功能机制。总之,DRT9-ncRNA复合物通过形成高度稳定的六聚体结构,使ncRNA紧密包裹DRT9蛋白,从而发挥独特的抗噬活性。这种结构特点可能是DRT9区别于其他逆转录酶的关键所在。
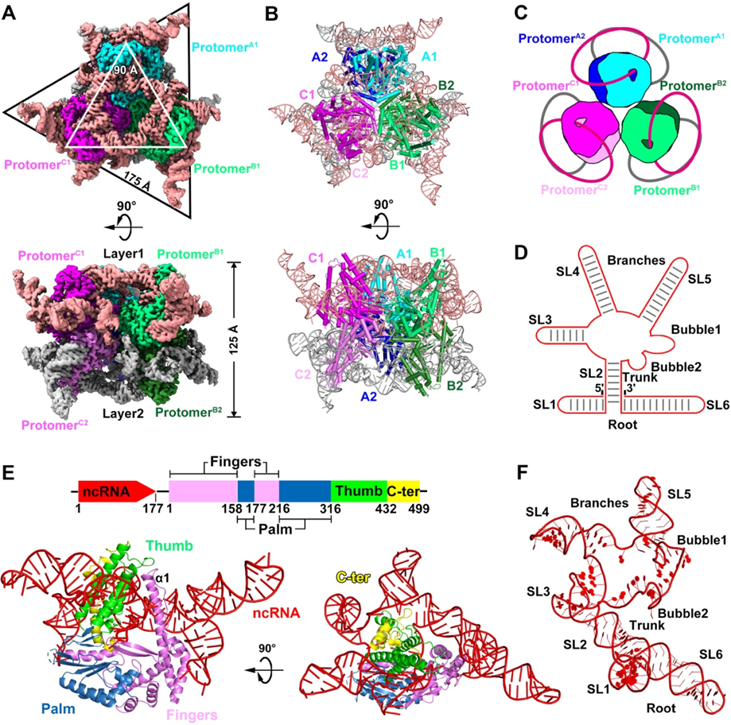
图2 DRT9-ncRNA六聚体复合体的整体结构
作者通过与已知RT结构(GsI-IIC RT)的比较分析,发现DRT9保留了经典的RT活性中心结构域,包括YADD催化位点、dNTP结合口袋、引物结合区等关键结构元件。对这些结构域进行突变实验,证实了它们对DRT9抗噬活性的关键作用。这表明DRT9采用与其他RT相似的反转录机制发挥功能。与此同时,DRT9也具有独特的结构特征,如指结构域中的延长α1螺旋,以及拇指结构域内的小α12螺旋和C端延长的4个α螺旋。这些独特结构要素能够与ncRNA紧密相互作用,诸如α1螺旋与拇指结构域的相互作用,形成一种"封闭"构象以包裹ncRNA。进一步实验证明,破坏这些独特的结构元件都会导致DRT9丧失抗噬活性,表明它们对DRT9-ncRNA复合物功能的关键性。此外,DRT9所结合的177nt ncRNA本身也具有特殊的二级结构,包括6个茎环,破坏这些结构也会影响DRT9的活性。综上所述,DRT9独特的结构特征及其与特定ncRNA的稳定复合是DRT9发挥高效抗噬功能的关键基础。
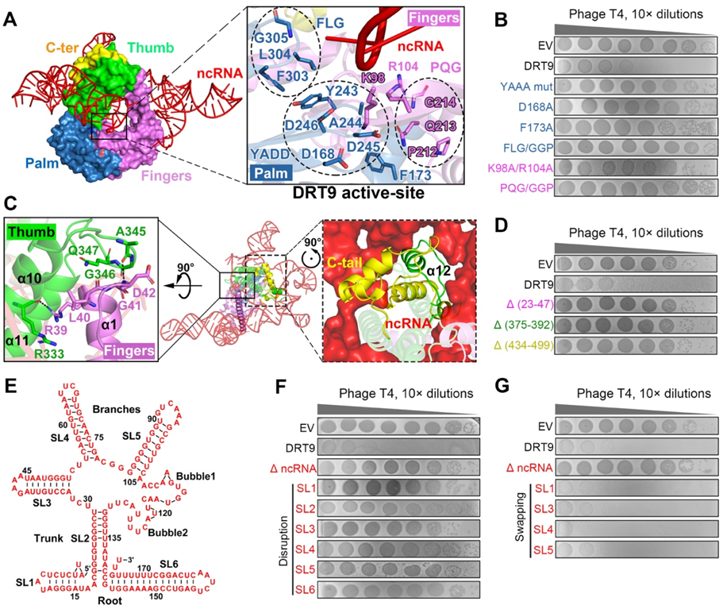
图3 DRT9和ncRNA的关键结构特征对抗噬菌体防御活性至关重要
作者通过凝胶过滤层析(SEC)可以观察到DRT9-ncRNA存在两种形式:寡聚体和单体。只有寡聚体具有cDNA合成活性,而单体则无活性。结构分析表明,DRT9-ncRNA复合物组装成一个六聚体结构,呈"三聚二聚体"配置。这个六聚体结构主要由两个层面的稳定作用维持:二聚体界面:包括指结构域的α3螺旋和掌状结构域的α4螺旋之间的氢键和盐桥作用;三聚体界面:涉及α9螺旋与β7-β8环之间的氢键作用。破坏这些关键界面上的氨基酸,如K127/H130/R131或Y254,都会导致六聚体的解离和cDNA合成活性的丧失,说明它们对复合物功能的关键性。此外,ncRNA中的一些结构元素,如SL5与bubble 1之间的G-C碱基对,也在维持六聚体结构中发挥重要作用。破坏这些ncRNA结构同样会导致复合物活性下降。综上所述,DRT9-ncRNA六聚体结构的形成和稳定,需要同时依赖于DRT9蛋白的寡聚化和ncRNA特有的结构特征,这是DRT9发挥抗噬活性的必要条件。
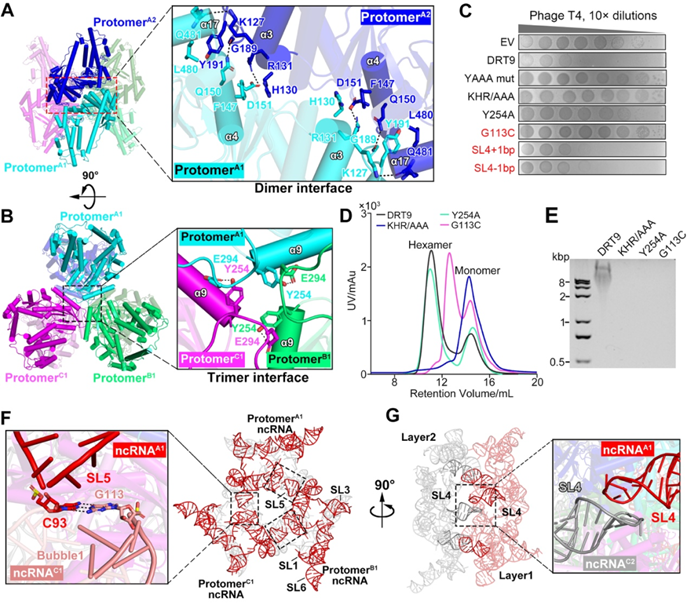
图4 DRT9-ncRNA的寡聚化是cDNA合成的必要条件
体外实验证实,DRT9-ncRNA复合物可合成大量的poly-A富集单链cDNA。体内实验也发现,在噬菌体感染过程中,细胞中存在大量的poly-A序列的cDNA产物进一步研究发现,尽管DRT9表达细胞中dATP水平下降,但仍高于逃逸性噬菌体感染的细胞,排除了通过耗竭dATP来抑制噬菌体复制的可能性。RNA测序分析显示,DRT9表达细胞中噬菌体DNA复制相关基因(特别是编码单链DNA结合蛋白SSB的基因)表达大量增加,但实际噬菌体DNA复制和释放仍受到抑制。进一步实验证明,过量表达噬菌体SSB蛋白可以部分抑制DRT9的抗噬活性,表明SSB的功能被干扰是DRT9抗噬的关键机制。体外实验证实,DRT9合成的poly-A富集单链cDNA可以直接结合噬菌体SSB蛋白,阻碍其参与噬菌体DNA复制。综上所述,DRT9系统合成大量无结构的poly-A cDNA,通过直接吸附和隔离噬菌体SSB蛋白,从而有效地抑制噬菌体的复制和扩散,发挥其强大的抗噬防御功能。
图5 DRT9合成的富含多聚A的长链ssDNA破坏噬菌体繁殖
在这项研究中,作者证明了DRT9与其上游的177-nt ncRNA结合,通过流产感染赋予抗噬菌体防御,特别是通过催化合成富含多聚A的长单链cDNA。此外,作者确定了DRT9-ncRNA六聚体复合物的2.62 Å冷冻电镜结构,为DRT9介导的富含多聚 A的cDNA合成提供了机制见解。基于结构分析,结合体内和体外生化研究,作者提出了DRT9介导的抗噬菌体防御机制模型。本研究提供了抗病毒免疫系统DRT9如何通过合成富含多聚A的长单链cDNA来防御噬菌体感染的机制见解,突出了DRT的多种抗噬菌体策略,揭示了逆转录酶的新功能维度,并为开发基于DRT9的生物技术工具奠定基础。
原文链接:https://www.science.org/doi/10.1126/science.ads4639
1、凡本网所有原始/编译文章及图片、图表的版权均属微生物安全与健康网所有,未经授权,禁止转载,如需转载,请联系取得授权后转载。
2、凡本网未注明"信息来源:(微生物安全与健康网)"的信息,均来源于网络,转载的目的在于传递更多的信息,仅供网友学习参考使用并不代表本网同意观点和对真实性负责,著作权及版权归原作者所有,转载无意侵犯版权,如有侵权,请速来函告知,我们将尽快处理。
3、转载请注明:文章转载自www.mbiosh.com
联系方式:020-87680942

