
原噬菌体介导的抗病毒防御与病毒反防御机制

为模型,揭示了温和噬菌体通过前噬菌体表达多种防御系统抵抗外源病毒感染的机制,并发现病毒进化出特异性反防御策略以突破宿主限制。&summary=&title=原噬菌体介导的抗病毒防御与病毒反防御机制&pics=https://mbiosh-prod.oss-cn-guangzhou.aliyuncs.com/upload/1751503990375/图片1.png&style=203&width=19&height=22)
1. 前噬菌体编码的多样化抗病毒防御系统
单亚基限制系统:Panchino噬菌体的gp28蛋白(类似I型限制修饰系统)可切割外源DNA,阻断病毒复制,但允许宿主细胞存活。异源排斥系统:Charlie噬菌体的gp32蛋白(膜定位)特异性阻止Che9c噬菌体的DNA注入,但对自身及近缘噬菌体无效。(p)ppGpp合成酶系统:Phrann和MMB噬菌体的gp29-gp30复合物通过合成警报素(p)ppGpp抑制宿主代谢,阻断病毒增殖,且依赖病毒早期基因激活。毒素-抗毒素(TA)系统:Xeno等噬菌体的TA系统可能通过诱导细胞休眠抑制病毒复制。

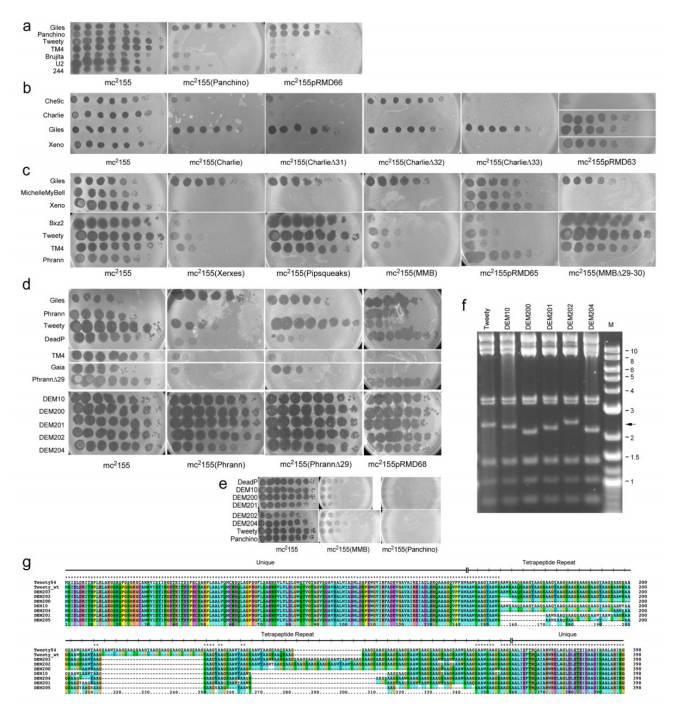
图1. N簇原噬菌体介导的防御的遗传学
2. 防御系统的特异性与宿主适应性
防御靶标高度特异:Panchino防御覆盖29种噬菌体,Charlie仅针对Che9c,而Phrann/MMB系统靶向Tweety等少数病毒。防御不依赖同源免疫:多数系统(如gp28、gp32、(p)ppGpp)与超感染免疫无关,属于异源防御。宿主存活优先:防御通常不导致细胞死亡,而是通过限制病毒增殖或抑制代谢实现宿主存活,与CRISPR等致死性防御不同。
3. 病毒的反防御进化机制
Tweety噬菌体的gp54蛋白:含高度重复的tetrapeptide序列,通过改变重复单元数量(±5-6个)获得反防御能力,特异性中和Phrann/MMB系统。基因突变逃逸:Tweety Δ54突变体通过gp56/gp57基因突变(如提前终止密码子)绕过防御,提示病毒早期基因参与防御识别。功能获得性突变:gp54的重复数变化并非失活,而是功能增强,表明病毒通过快速重组进化出适应性反防御策略。
4. 防御与反防御的协同进化
前噬菌体防御系统与病毒反防御形成动态博弈:Phrann/MMB系统通过(p)ppGpp抑制病毒,但Tweety gp54可阻断该通路;Charlie gp32排斥Che9c,但Che9c尚未发现反防御机制;Panchino gp28限制DNA,但病毒可能通过甲基化修饰逃逸。防御系统的多样性(限制、排斥、代谢抑制)与病毒反防御的特异性(如gp54仅针对(p)ppGpp系统)共同驱动宿主-病毒共进化。
5. 生态与进化意义
前噬菌体防御是细菌应对病毒压力的重要策略,尤其在分枝杆菌等慢性感染病原体中,可能增强宿主环境适应性。病毒反防御机制(如gp54的可调重复序列)揭示了病毒通过基因组可塑性快速适应宿主防御的进化潜力。防御与反防御的相互作用为理解微生物群落中宿主-病毒平衡、基因水平转移及耐药性进化提供了新视角。
结论:本研究首次系统揭示了温和噬菌体通过前噬菌体编码多种防御系统抵抗病毒感染的机制,并阐明了病毒通过特异性反防御策略突破宿主限制的进化路径。这些发现不仅拓展了病毒-宿主互作的理论框架,也为利用噬菌体防御系统开发新型抗菌疗法提供了潜在靶点。
参考来源:
10.1038/nmicrobiol.2016.251
1、凡本网所有原始/编译文章及图片、图表的版权均属微生物安全与健康网所有,未经授权,禁止转载,如需转载,请联系取得授权后转载。
2、凡本网未注明"信息来源:(微生物安全与健康网)"的信息,均来源于网络,转载的目的在于传递更多的信息,仅供网友学习参考使用并不代表本网同意观点和对真实性负责,著作权及版权归原作者所有,转载无意侵犯版权,如有侵权,请速来函告知,我们将尽快处理。
3、转载请注明:文章转载自www.mbiosh.com
联系方式:020-87680942

