
噬菌体进化出对黏膜表面的增强黏附能力

系统,首次实证了噬菌体(T4)不仅与其细菌宿主共同进化,还能直接适应哺乳动物肠道黏膜环境,形成三者之间的三方共进化关系。&summary=&title=噬菌体进化出对黏膜表面的增强黏附能力&pics=https://mbiosh-prod.oss-cn-guangzhou.aliyuncs.com/upload/1752656366295/图片1.png&style=203&width=19&height=22)
一、噬菌体与细菌在肠道芯片中稳定共存
在模拟肠道黏膜的三维微流控系统中,T4噬菌体与大肠杆菌(E. coli)在黏膜层中实现了稳定共存。尽管不同实验重复之间的种群动态存在显著差异,但噬菌体和细菌均能在24小时内持续存在。这一现象与传统试管环境中细菌频繁灭绝的情况形成鲜明对比,说明黏膜层的空间异质性为细菌提供了“避难所”,从而缓解了噬菌体的捕食压力,维持了种群的动态平衡。

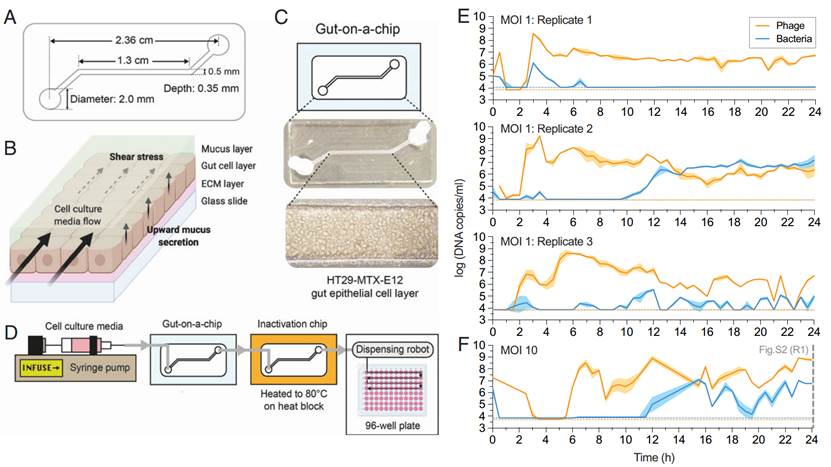
图1. 芯片上的肠道支持粘液环境中的噬菌体-细菌共培养
二、噬菌体在黏膜环境中出现适应性突变
经过五轮连续传代实验,仅在肠道芯片中培养的噬菌体种群中出现了高频突变,而在试管对照组中未观察到类似趋势。最主要的突变包括:Hoc蛋白D246N突变:位于噬菌体外壳蛋白Hoc的第三个Ig-like结构域,改变了其与岩藻糖基化黏蛋白聚糖的结合能力;goF基因Δ21bp缺失:导致七个氨基酸缺失,可能影响RNA转录调控,但其表型效应尚不明确。这两个突变通过基因重组整合到同一基因组中,形成了双突变株(D246N::ΔgoF),并在种群中迅速上升,表明其在黏膜环境中具有选择优势。
三、高感染复数(MOI)促进噬菌体重组
实验发现,肠道芯片中噬菌体与细菌的比例(MOI)较高,促进了噬菌体之间的共感染与基因重组。通过构建突变株与野生株的混合感染实验,证实高MOI条件(MOI = 10)下的重组频率显著高于低MOI(MOI = 0.1)。这表明,黏膜环境中的高噬菌体密度为有益突变的重组与固定提供了机制基础。
四、突变型噬菌体在黏膜环境中具有竞争优势
将D246N::ΔgoF突变株与祖先型噬菌体在肠道芯片中进行竞争实验,结果显示突变株在10次重复实验中有3次成功取代祖先型,并在一次实验中达到固定,平均选择系数为正(s ≈ 0.6)。尽管整体差异未达统计学显著水平,但趋势明确,表明该突变在黏膜环境中具有温和但稳定的适应性优势。
五、Hoc突变改变噬菌体与岩藻糖基化聚糖的亲和力
通过糖微阵列和表面等离子共振(SPR)实验,研究发现D246N突变显著降低了Hoc蛋白与岩藻糖基化聚糖(如Lacto-N-fucopentaose I)之间的结合亲和力。进一步通过肠道芯片中的滞留-洗脱实验验证,在岩藻糖基化聚糖存在下,D246N突变株比野生型更不易被洗脱,表现出更强的黏膜滞留能力。这一结果说明,降低对岩藻糖基化聚糖的结合反而有助于噬菌体避免被可溶性聚糖“捕获”,从而增强其在黏膜中的持续存在。
六、噬菌体适应性进化反映宿主个体化特征
研究强调,人类肠道黏膜的岩藻糖基化模式具有高度个体差异性,主要由宿主FUT2基因型决定。噬菌体通过其表面蛋白(如Hoc)快速适应这些个体特异的糖基化结构,提示其在人肠道生态系统中可能参与形成个体特异的病毒组(virome)结构。这种“宿主-病毒共塑”机制为理解个性化微生物组提供了新视角。
七、研究意义与未来方向
本研究首次提供了噬菌体在哺乳动物黏膜环境中适应性进化的实验证据,突破了传统“噬菌体-细菌”二元进化框架,提出“噬菌体-细菌-宿主”三方共进化的可能性。研究结果不仅拓展了对肠道病毒生态的理解,也为未来基于噬菌体的精准医疗提供了理论基础。未来研究可进一步拓展至多噬菌体、多菌株及多宿主糖型背景下的复杂共进化网络,并探索其在疾病干预中的潜在应用。
参考来源:
10.1073/pnas.2116197119
1、凡本网所有原始/编译文章及图片、图表的版权均属微生物安全与健康网所有,未经授权,禁止转载,如需转载,请联系取得授权后转载。
2、凡本网未注明"信息来源:(微生物安全与健康网)"的信息,均来源于网络,转载的目的在于传递更多的信息,仅供网友学习参考使用并不代表本网同意观点和对真实性负责,著作权及版权归原作者所有,转载无意侵犯版权,如有侵权,请速来函告知,我们将尽快处理。
3、转载请注明:文章转载自www.mbiosh.com
联系方式:020-87680942

