
转录因子AreA对灵芝胞外和胞内多糖含量的氮依赖性调控


灵芝作为东南亚有名的大型食药用菌,含有多种有效活性成分,包括多糖,三萜等等对人体有益的代谢物。因此,如何提高灵芝活性成分含量成为人们关注的焦点
氮源是影响真菌生长发育和代谢的重要营养物质之一。真菌优先利用首选氮源,如易于同化的氨和谷氨酰胺。只有在首选氮源耗尽后,他们才会利用非首选氮源。当首选氮源可用时,这种现象被称为氮分解代谢抑制[1]。同时,在多项研究中,氮源的增加能有效提高多个物种的多糖含量 [2-4]。因此,通过调控氮源的合成途径相关基因从而调控多糖含量可能是一种可行的方法。
2025年4月南京农业大学朱静团队在国际杂志Microbiological Research中发表了《Nitrogen-dependent regulation of extracellular and intracellular polysaccharide content in Ganoderma lucidum via the transcription factor AreA》[5],阐明了氮源对多糖合成的影响,为提高灵芝多糖含量的策略提供了有价值的见解。

1 不同氮源下灵芝的转录组学特征
研究人员对灵芝基因组进行了转录组分析以评估相关基因表达模式。与氨(首选氮源)相比,硝酸盐(非首选氮源)处理下523个基因上调,539个基因下调(图1A)。在这些基因中,下调的基因主要参与包括类固醇生物合成、甾醇生物合成和次生代谢,以及氧化还原酶活性和催化活性等生物过程以及与碳代谢和氨基酸生物合成相关的途径;上调的基因主要参与与质膜部分、膜的内在成分、跨膜转运蛋白活性和离子跨膜转运酶活性相关以及与多糖合成、几丁质合成和氨基酸跨膜转运相关的生物过程以及戊糖和葡萄糖醛酸的相互转化、糖酵解/糖异生以及氨基糖和核苷酸糖代谢等途径(图1B-E)。
在转录分析中,研究人员还对影响多糖合成的基因表达水平进行分析,发现在硝酸盐的处理下,这些基因的表达水平上升50-90%不等(图2A)。而调控灵芝胞内多糖和胞外多糖,细胞壁多糖合成基因也有不同程度的表达上调(图2B-C)。结果表明,非首选氮源的培养条件显著诱导了灵芝多糖合成相关基因的表达。
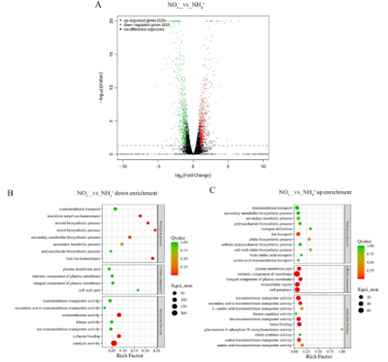
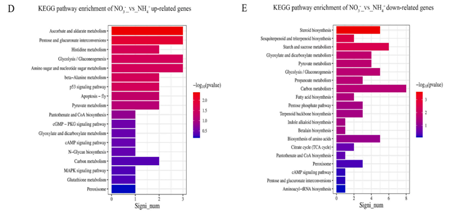
图1 不同氮源(氨或硝酸盐)下的转录组分析
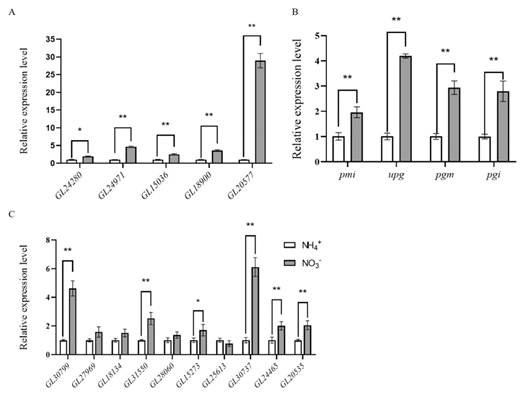
图2 多糖合成中基因的表达分析
2 氮源对灵芝多糖合成的影响
研究人员对不同培养时间的菌丝生物量以及胞外多糖(EPS)和胞内多糖(IPS)的含量进行了检测,发现在转移到氨态氮条件后的五天内,生物量在前72小时内迅速增加,但在接下来的两天内几乎停止增加。相比之下,在硝酸盐条件下,前48小时的生长相对缓慢,随后增长更快(图3A)。与硝态氮硝酸盐相比,氨处理下EPS的含量始终较高(图3B)。然而,硝酸盐条件下的IPS含量始终高于氨条件下的含量(图3C)。除此以外,硝酸盐条件下细胞壁多糖甲壳质和葡聚糖含量均显著高于氨条件下(图3D-E)。结果表明,使用非首选氮源(硝酸盐)不利于EPS的积累,但会导致灵芝中IPS含量的增加。
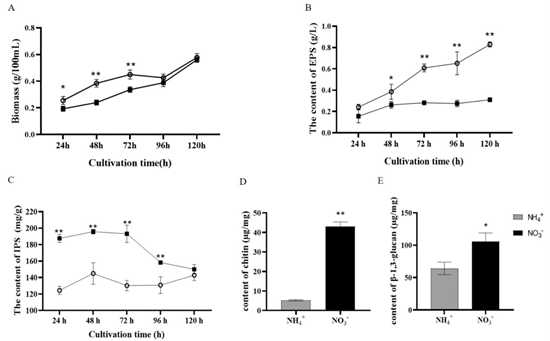
图3 不同氮源对灵芝多糖含量的影响
3 不同氮源下芳烃对EPS和IPS合成的影响
转录因子AreA在氮分解代谢抑制中起着重要作用[1,6],但其在调节多糖合成中的作用相对较少。为确定氮源对IPS和EPS合成的影响是否受到转录因子AreA的影响。在氨或硝酸盐条件下测量野生型(WT)和areA沉默菌株中IPS和EPS的含量。在氨处理条件下,areA沉默菌株和WT之间的IPS和EPS含量没有显著差异。然而,在硝酸盐条件下,与WT相比,areA沉默菌株中的IPS含量显著降低了约24%,而EPS含量显著增加了约33%(图4A-B)。
在硝酸盐条件下,WT和areA沉默菌株的多糖合成相关基因pmi表达没有显著变化。在氨条件下,areA沉默菌株中pmi的表达水平增加了40-60%(图4C)。在氨条件下,areA沉默菌株中pgi、ugp和pgm的表达没有显著变化。然而,在硝酸盐条件下,它们显著减少了约66%(图4D-F)。结果表明,AreA参与调节灵芝中IPS和EPS的合成生物过程。
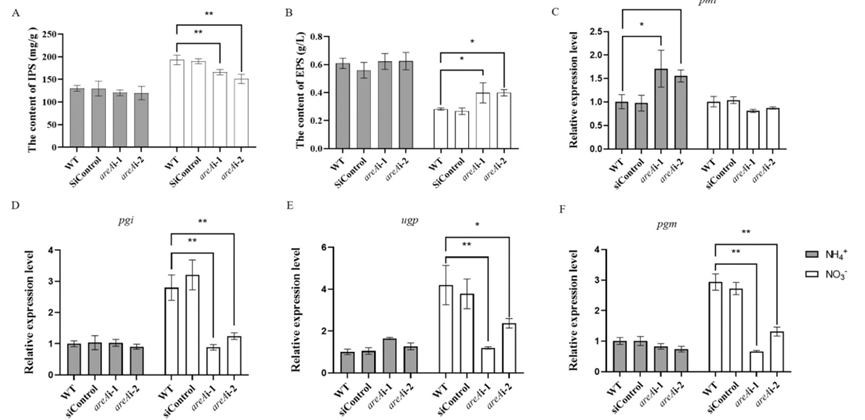
图4 不同氮源下WT和areA沉默菌株IPS和EPS含量及多糖合成相关基因表达的分析
4 不同氮源下AreA对甲壳素和β-1,3-葡聚糖合成的影响
研究人员研究了氮源对细胞壁多糖甲壳质和β-1,3-葡聚糖合成的影响是否受到芳烃的影响。
在氨或硝酸盐条件下测量了WT和areA沉默菌株中几丁质的含量和几丁质合酶基因的表达(图5A和C-D)。在氨态条件下,对照菌株中甲壳质的含量比硝酸盐条件低约30-50%,沉默areA后,几丁质含量或大多数几丁质合酶基因的表达没有显著变化。然而,在硝酸盐条件下,与野生型相比,areA沉默菌株中的几丁质含量显著降低了约20%,几丁质合酶基因的表达显著降低了25%-80%。
在沉默 areA 后,无论是在铵态氮还是硝态氮条件下,β-1,3-葡聚糖的含量均显著降低了 20% - 25%,但在硝态氮条件下其含量仍高于铵态氮条件(图 5B)。在铵态氮条件下,沉默 areA 并未显著影响β-葡聚糖合酶基因(Gl24465)的表达,而葡聚糖合酶基因(Gl20535)的表达则显著降低了 50%。在硝态氮条件下,两个β-葡聚糖合酶基因的表达均显著降低了 55% - 60%(图 5C - D)。总之,areA 促进了在非首选氮源条件下几丁质和β-葡聚糖的积累。
结果表明,AreA在非首选氮源下促进甲壳质和β-葡聚糖的积累。
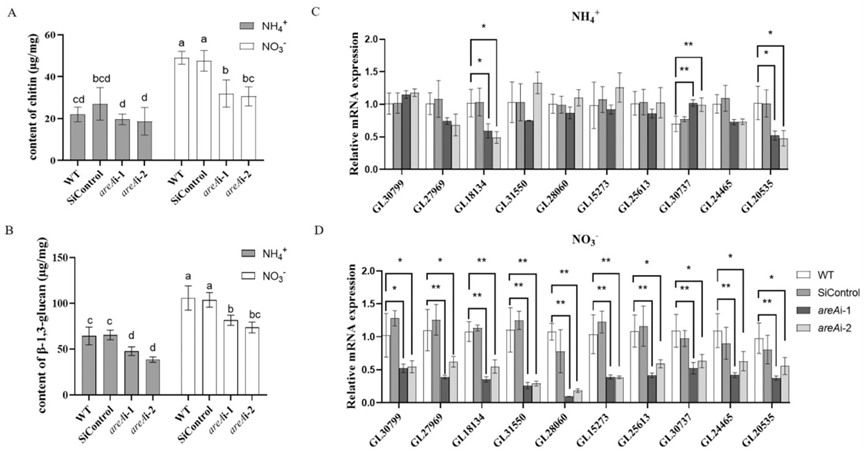
图5 不同氮源下WT和areA沉默菌株几丁质和β-1,3-葡聚糖含量及相关基因表达分析
5 AreA影响细胞壁的厚度和完整性
为了研究AreA对细胞壁的影响,研究人员使用透射电子显微镜观察了菌丝体的细胞壁(图6A-B)。在氨条件下,与WT菌株相比,areA沉默菌株的细胞壁厚度没有显著变化,而在硝酸盐条件下,细胞壁厚度减少了约23%。
为了确定转录因子AreA在细胞壁完整性中的作用,我们比较了WT和areA沉默菌株在添加细胞壁干扰剂的培养基上的生长情况。在areA沉默后,其对刚果红(CR)和Calcofluor White(CFW)的敏感性提高了20%-30%(图6C-D),而对CR和CFW。Slt2是细胞壁完整性(CWI)通路中的一种激酶,与WT相比,在areA沉默后,其转录水平显著降低。Swi6和Rlm1是CWI通路下游的转录因子。,沉默areA后,areAi-1和areAi-2菌株中swi6的转录水平分别提高了29%和50%,而rlm1的转录水平降低了。此外,我们检测了钙信号通路中的关键转录因子Crz1,发现其转录水平在areAi-1和areAi-2菌株中分别显著提高了65%和140%。
结果表明,AreA影响细胞壁的完整性,并与细胞壁完整性相关的途径有复杂的相互作用。
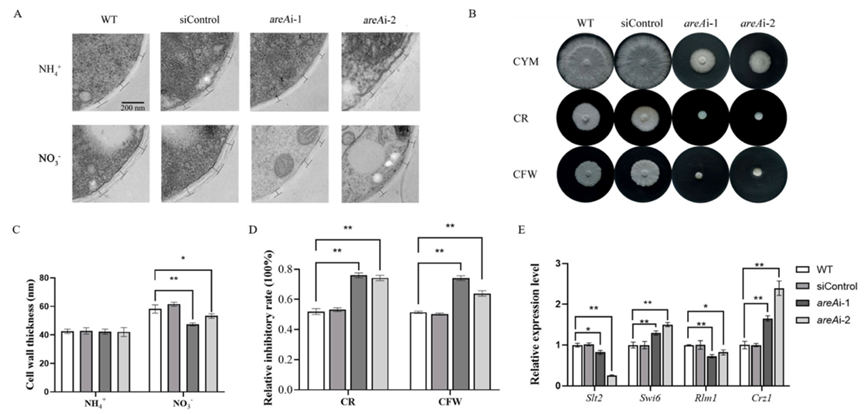
图6 AreA调节灵芝细胞壁的结构和完整性
总而言之,该研究阐明了不同氮源下灵芝的转录组学特征、氮源对灵芝多糖合成的影响、不同氮源下芳烃对EPS和IPS合成的影响与AreA对甲壳素和β-1,3-葡聚糖合成的影响以及AreA对灵芝细胞壁的厚度和完整性影响进行了相关探究,为灵芝多糖合成途径研究提供了坚实的理论基础。
参考文献
[1] Wong Koon, H., Hynes Michael, J., Davis Meryl, A., 2008. Recent advances in nitrogen regulation: a comparison between Saccharomyces cerevisiae and filamentous fungi. Eukaryot. Cell 7 (6), 917–925. https://doi.org/10.1128/ec.00076-08.
[2] Wang, W., Liu, Y., Geng, L.L., Yang, Z., Chen, Y.F., 2011. The effect of nitrogen concentration and its interaction with light intensity on population dynamics and polysaccharide content of Microcystis aeruginosa. J. Freshw. Ecol. 26 (2), 241–248. https://doi.org/10.1080/02705060.2011.555205.
[3] Liu, X.H., Ou, H.P., Yang, R.Z., Zhang, R.H., Yang, L.T., Li, Y.R., 2016. Change of polysaccharide content and its relation with nitrogen level in sugarcane leaves under nitrogen-fixing bacteria inoculation and nitrogen supply. Sugar Tech 18 (1), 87–92. https://doi.org/10.1007/s12355-015-0368-z.
[4] Liang, X., An, W., Li, Y., Qin, X., Zhao, J., Su, S., 2024. Effects of different nitrogen application rates and picking batches on the nutritional components of Lycium barbarum L. fruits. Front. Plant Sci. 15, 1355832. https://doi.org/10.3389/ fpls.2024.1355832.
[5] Guo, X., Yang, Y., Li, Y., Chen, B., Li, H., Zhang, C., Ma, J., Zhao, M., Zhu, J., 2025. Nitrogen-dependent regulation of extracellular and intracellular polysaccharide content in Ganoderma lucidum via the transcription factor AreA. Microbiological research, 297, 128197. https://doi.org/10.1016/j.micres.2025.128197
[6] Giese, H., Sondergaard, T. E., & Sørensen, J. L. (2013). The AreA transcription factor in Fusarium graminearum regulates the use of some nonpreferred nitrogen sources and secondary metabolite production. Fungal biology, 117(11-12), 814–821. https://doi.org/10.1016/j.funbio.2013.10.006
1、凡本网所有原始/编译文章及图片、图表的版权均属微生物安全与健康网所有,未经授权,禁止转载,如需转载,请联系取得授权后转载。
2、凡本网未注明"信息来源:(微生物安全与健康网)"的信息,均来源于网络,转载的目的在于传递更多的信息,仅供网友学习参考使用并不代表本网同意观点和对真实性负责,著作权及版权归原作者所有,转载无意侵犯版权,如有侵权,请速来函告知,我们将尽快处理。
3、转载请注明:文章转载自www.mbiosh.com
联系方式:020-87680942

