
胖孩子的肠道危机:哪些细菌在推动脂肪肝变“肝炎”?

一步步变成更严重的“脂肪性肝炎”(MASH)。研究找到了关键的细菌种类和代谢变化,不仅能提前预警肝病进展,还提醒我们:调整孩子的饮食,可能从源头拦住肝病恶化。&summary=&title=胖孩子的肠道危机:哪些细菌在推动脂肪肝变“肝炎”?&pics=https://mbiosh-prod.oss-cn-guangzhou.aliyuncs.com/upload/1749732805231/图片1.jpg&style=203&width=19&height=22)
MASLD是儿童和青少年中最常见的慢性肝病原因,全球患病率在5%到8%之间。它可能进展为MASH,进而导致肝硬化和终末期肝病。MASLD的定义是超过5%的肝细胞中出现脂滴的病理性积累,并且至少满足一个心脏代谢标准,如糖调节受损或肥胖等。MASH则以肝内炎症和细胞损伤为特征。肠道微生物组是一个复杂的生态系统,与宿主共生,其组成变化会影响免疫发育、黏膜保护和代谢功能。近年来研究表明,肠道菌群的组成和功能异常可能使个体易患甚至加剧MASLD的发展。随着16S-rRNA测序和宏基因组测序技术的发展,研究发现肠道微生物群的组成变化与MASLD和MASH的发展有关。例如,MASLD儿童中拟杆菌门(Bacteroidota)减少和厚壁菌门(Firmicutes)增加,这些变化甚至可以在婴儿期早期观察到。此外,肠道微生物群的特定组成变化还可以预测成人队列中MASH疾病的进程。然而,目前缺乏对使用宏基因组学对肥胖和相关MASLD或MASH的儿童和青少年中肠道微生物组成和代谢功能进行大规模队列研究的系统性分析。基于此,《Gut Microbes》近期发表的一项研究采用鸟枪法宏基因组测序,整合9项国际研究及本地队列共353例样本(健康对照、肥胖无MASLD、MASLD、MASH儿童),并纳入163例成人MASH样本作为对照,通过机器学习算法(XGBoost、随机森林)系统解析如下2个关键科学问题:(1)儿童肠道菌群组成和代谢通路如何随MASLD严重程度(从单纯肥胖到MASH)发生改变;(2)儿童与成人MASH患者的肠道菌群差异是否由年龄相关代谢环境驱动,能否通过菌群特征实现疾病阶段和年龄组的精准区分。
研究结果显示,与健康儿童相比,MASLD和MASH儿童的肠道微生物组的α多样性和β多样性均显著不同。具体而言,MASH患者的微生物观测丰富度显著降低,而Shannon多样性无显著差异。在物种水平上,Faecalibacterium prausnitzii和Alistipes putredinis在肥胖、MASLD和MASH组之间的相对丰度存在显著差异。此外,机器学习模型能够准确区分肥胖与MASLD(AUROC为87%),以及MASLD与MASH(AUROC为89%),图1。这些发现表明,肠道微生物组的组成随着MASLD向MASH的进展而发生显著变化,且这些变化可以作为潜在的生物标志物用于疾病的诊断和进展评估。作者进一步研究了代谢功能,发现MASLD患者与肥胖患者相比代谢通路变化较小,而MASLD与MASH患者之间的代谢通路变化更为显著。特别是,“nylon-6 oligomer degradation”通路在MASLD患者中显著高于MASH患者。基于代谢通路丰度的XGBoost模型能够以81%的AUROC区分MASLD和肥胖,以88%的AUROC区分MASH和MASLD,相关结果见图2。这些代谢通路的变化反映了肠道微生物群在疾病进展中的功能重塑。
在儿童与成人MASH患者的比较中,研究发现两者的肠道微生物组存在显著差异,机器学习模型能够以97%的AUROC区分儿童和成人MASH患者,图3。这提示菌群差异与年龄相关代谢环境有关。这一发现具有重要的治疗意义。首先,肠道微生物组的变化可以作为粪便生物标志物,提示饮食干预对MASLD/MASH进展的潜在调控作用。其次,由于儿童和成人的肠道微生物组存在显著差异,针对儿童的治疗策略可能需要特别考虑其肠道微生物组的特性。例如,通过调整饮食习惯或补充特定的益生菌,可能有助于恢复儿童肠道微生物组的健康状态,从而减轻MASLD和MASH的严重程度。此外,考虑到儿童的肠道微生物组在发育过程中具有较高的可塑性,早期干预可能更为有效。
综上所述,本研究揭示了肠道菌群随MASLD进展的动态变化规律,证实了其作为无创诊断标志物的潜力,并强调了儿童与成人菌群特征的年龄特异性。未来研究可进一步探索菌群-代谢互作机制,以及通过饮食或微生态制剂干预疾病的可能性。
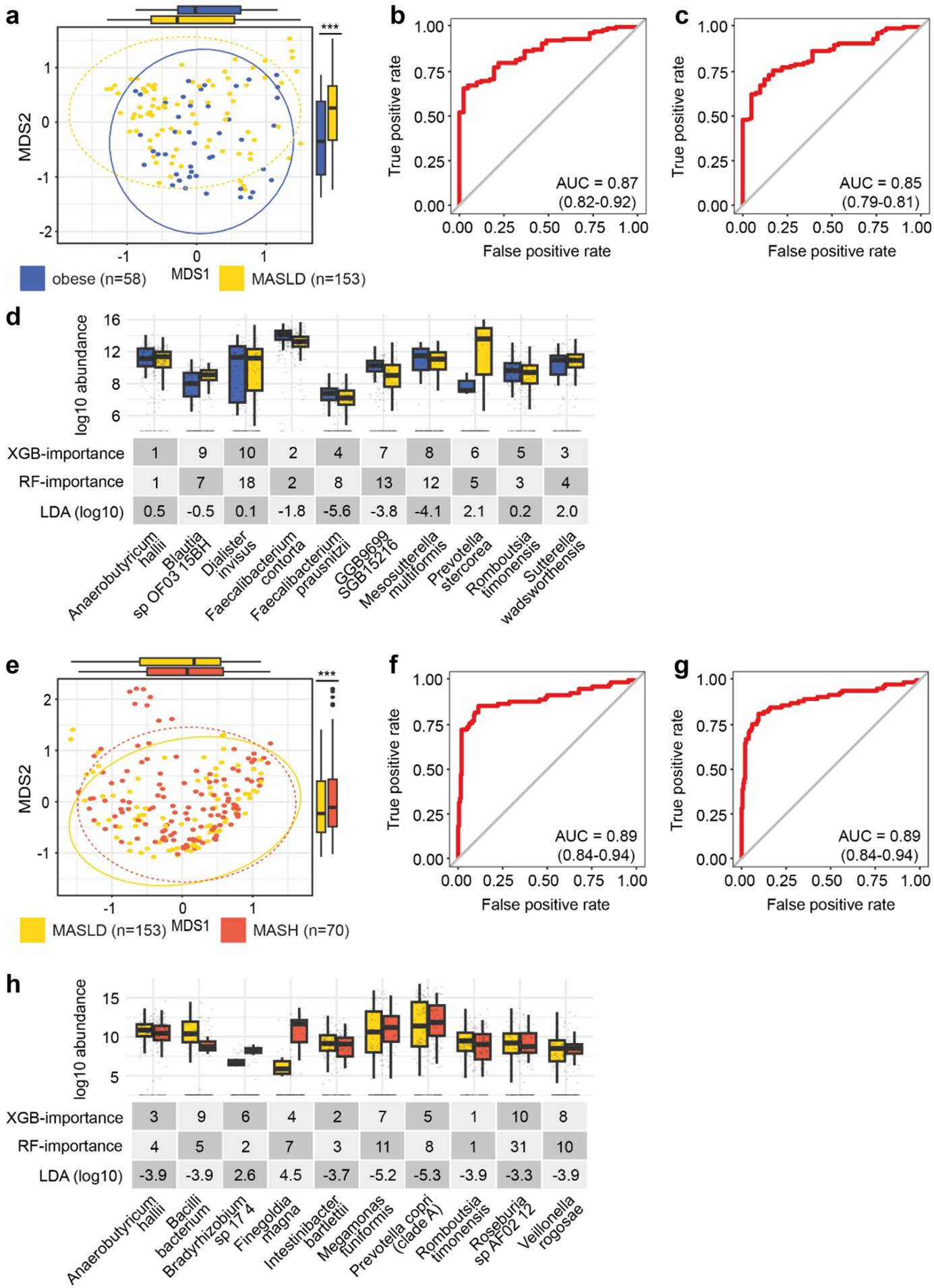
图1 群体多样性与疾病状态预测在MASLD和MASH中的表现:(a-e)通过主坐标分析(MDS)和Bray-Curtis距离度量,比较肥胖和MASLD(a)、MASLD和MASH(e)患者的肠道微生物组的β多样性。p<0.001。(b)XGBoost模型区分肥胖患者是否患有MASLD的AUROC为87%(CI0.82–0.92)。(c)随机森林模型区分肥胖患者是否患有MASLD的AUROC为85%(CI0.79–0.81)。(d)通过监督机器学习识别区分肥胖和MASLD患者的前十个关键微生物特征。(f)使用XGBoost模型基于肠道微生物组的组成预测MASLD患者是否进展为MASH,AUROC为89%(CI:0.84–0.94)。(g)使用随机森林模型基于肠道微生物组的组成预测MASLD患者是否进展为MASH,AUROC为89%(CI:0.84–0.94)。(h)通过监督机器学习识别区分MASLD和MASH患者的前十个关键微生物特征。
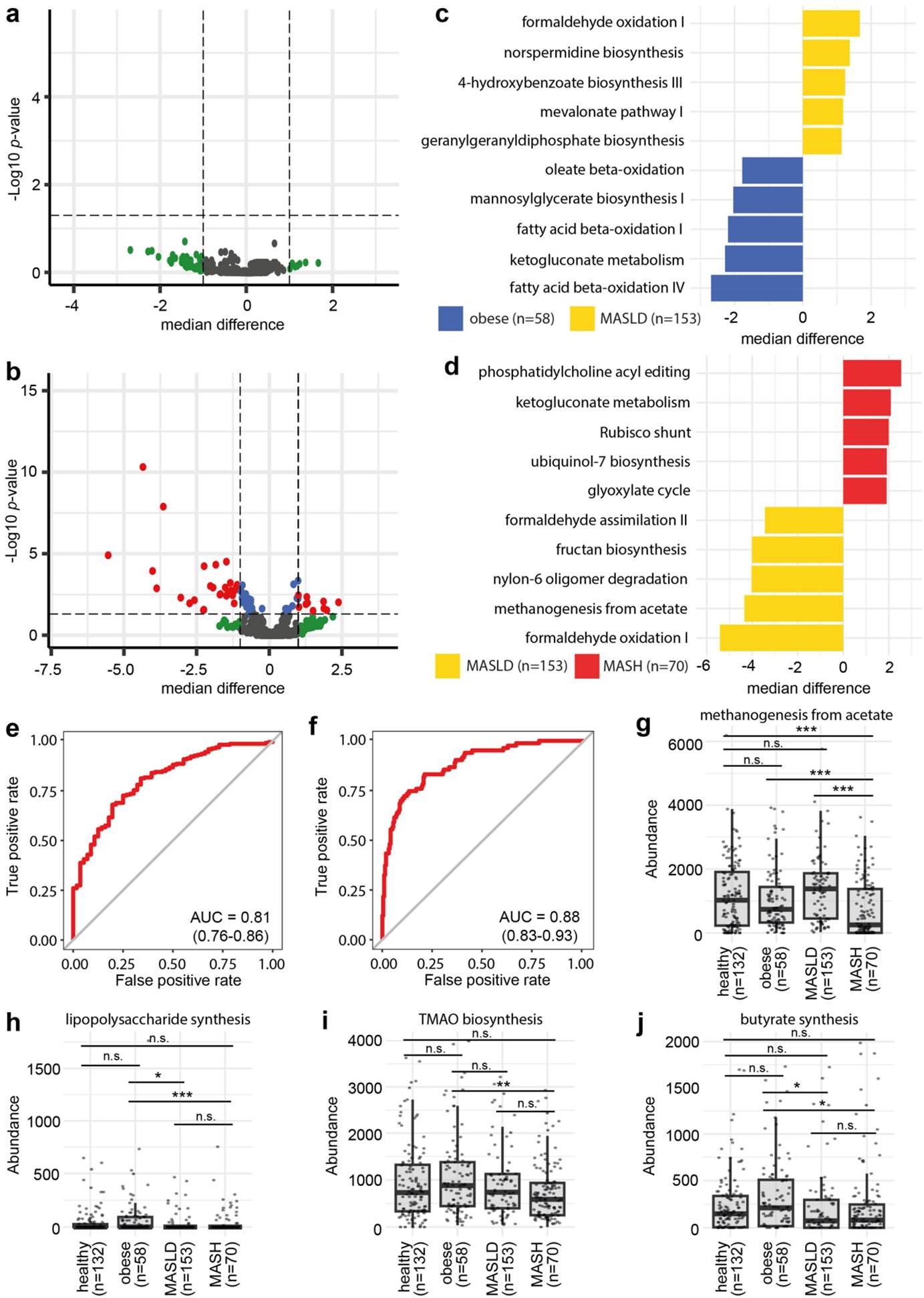
图2 (a)肥胖与MASLD患者代谢通路差异的火山图,未发现显著变化的代谢通路(log10(p)>1.3),表明肥胖和MASLD患者在代谢通路水平上的差异较小。(b)MASLD与MASH患者代谢通路差异的火山图,发现多个显著变化的代谢通路,例如“nylon-6 oligomer degradation”在MASLD患者中显著更高(log10(p)<-3)。(c)肥胖与MASLD患者代谢通路差异的发散条形图。(d)MASLD与MASH患者代谢通路差异的发散条形图。(e)基于代谢通路丰度的XGBoost模型预测MASLD,AUROC为81%(CI:0.76–0.86)。(f)基于代谢通路丰度的XGBoost模型预测MASH,AUROC为88%(CI:0.83–0.93)。(g)甲烷生成从乙酸通路的点箱图,甲烷生成通路的丰度差异可以作为区分MASLD和MASH的生物标志物。(h)LPS合成通路的点箱图,LPS合成通路在MASLD和MASH患者中显著降低,表明其可能与疾病的炎症反应有关。(i)TMAO生物合成通路的点箱图,TMAO生物合成通路在MASH患者中显著降低,表明其可能与疾病的进展有关。(j)丁酸合成通路的点箱图,丁酸合成通路在MASLD和MASH患者中显著降低,表明其可能与疾病的炎症和代谢功能障碍有关。
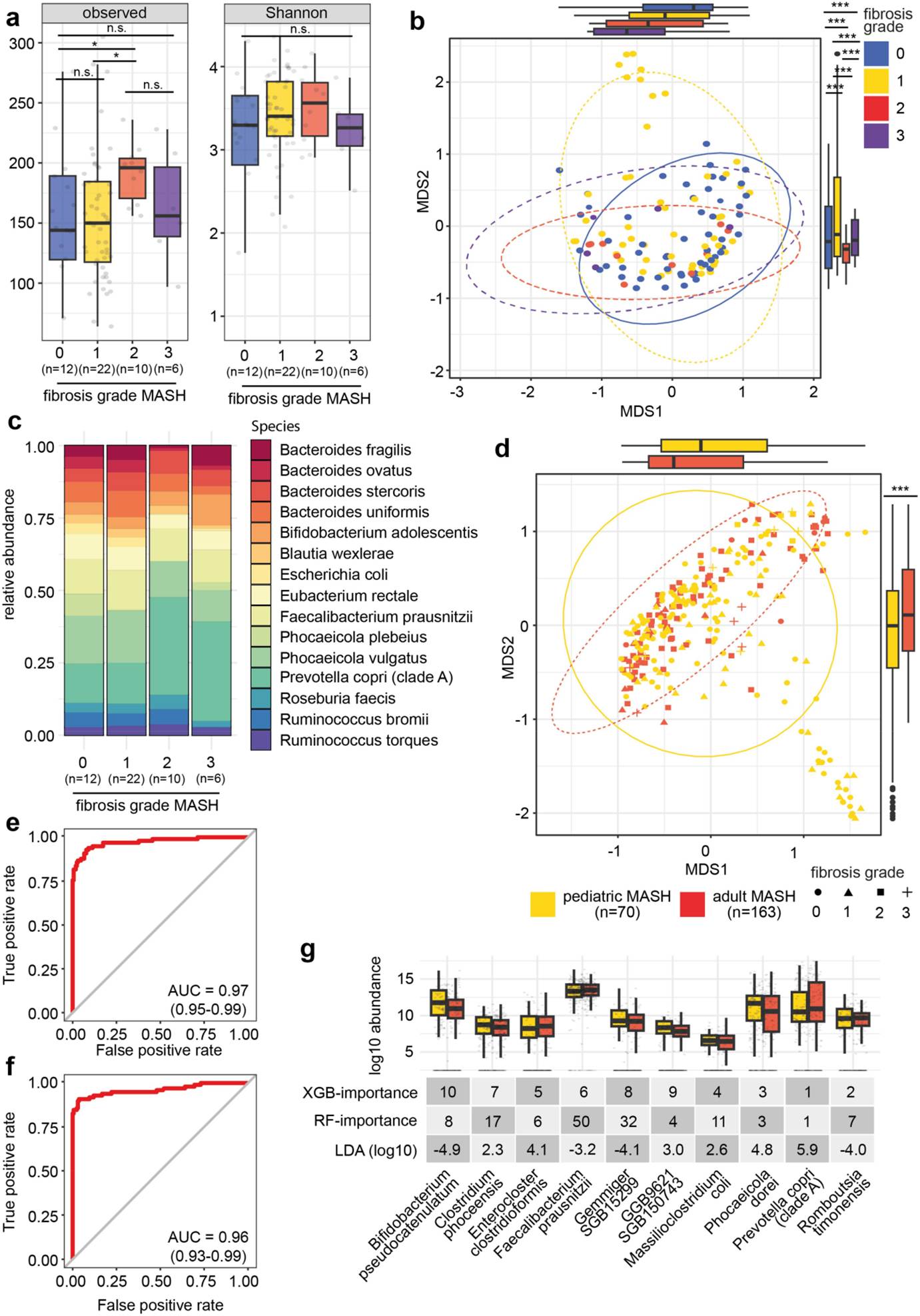
图3 肠道微生物动态变化与肝纤维化的关系。(a)肠道微生物丰富度在MASH患者中随肝纤维化程度增加而降低(p < 0.001)。(b)通过MDS和Bray-Curtis距离度量,分析不同肝纤维化程度的MASH患者的β多样性。不同肝纤维化程度的患者之间β多样性存在显著差异(p < 0.001)。(c)不同肝纤维化程度的MASH患者中前20个最丰富物种的相对丰度变化。某些物种(如Bifidobacterium adolescentis和Prevotella copri)的丰度随肝纤维化程度的变化而显著变化。(d)通过MDS和Bray-Curtis距离度量,比较儿童和成人MASH患者的β多样性。儿童和成人MASH患者的β多样性存在显著差异(p < 0.001)。(e)使用XGBoost模型基于肠道微生物组的组成区分儿童和成人MASH患者,AUROC为97%(CI 0.95–0.99)。(f)使用随机森林模型基于肠道微生物组的组成区分儿童和成人MASH患者,AUROC为96%(CI 0.93–0.99)。(g)通过监督机器学习识别区分儿童和成人MASH患者的前十个关键微生物特征。
原文链接:https://doi.org/10.1080/19490976.2025.2508951
1、凡本网所有原始/编译文章及图片、图表的版权均属微生物安全与健康网所有,未经授权,禁止转载,如需转载,请联系取得授权后转载。
2、凡本网未注明"信息来源:(微生物安全与健康网)"的信息,均来源于网络,转载的目的在于传递更多的信息,仅供网友学习参考使用并不代表本网同意观点和对真实性负责,著作权及版权归原作者所有,转载无意侵犯版权,如有侵权,请速来函告知,我们将尽快处理。
3、转载请注明:文章转载自www.mbiosh.com
联系方式:020-87680942

