
南京农业大学赵明文教授团队在AEM杂志发表亚精胺增强灵芝耐热性的最新进展


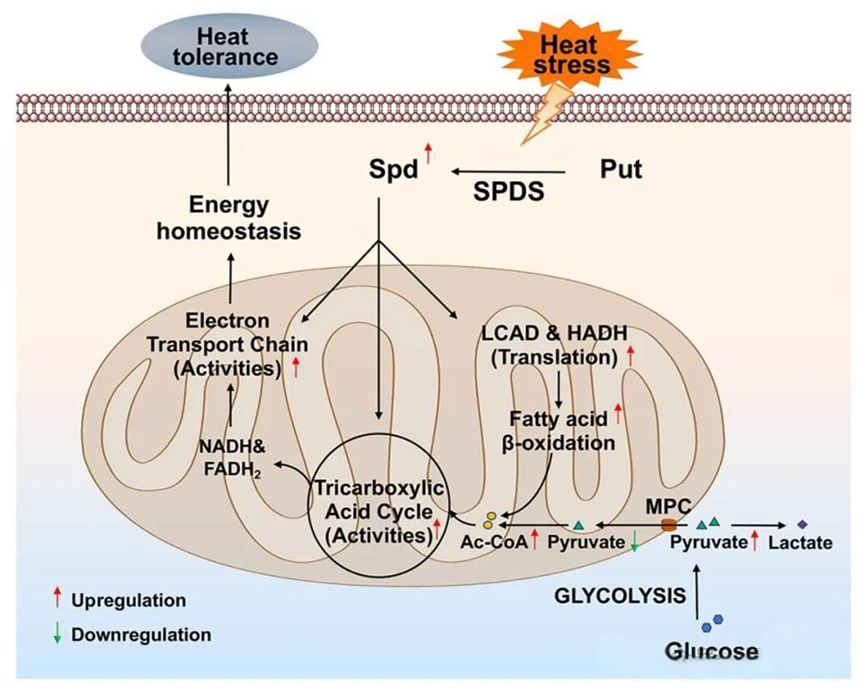
给大家分享南京农业大学赵明文教授团队的最新研究进展,关于亚精胺增强灵芝的耐热性。灵芝是我国著名的食药用真菌,是传统天然生物活性成分的最佳来源之一。前期研究发现热胁迫通过诱导亚精胺合成促进灵芝酸的积累(Han et al., Environmental Microbiology, 2022),但是亚精胺在灵芝抵抗热胁迫中的作用尚未可知。亚精胺是一种重要的多聚阳离子,参与了生物体内多种非生物胁迫响应。该团队研究发现亚精胺通过促进脂肪酸β-氧化驱动的线粒体呼吸增强灵芝的耐热性,成果在美国微生物学会旗下期刊《Applied and Environmental Microbiology》上发表,题为“Spermidine enhances the heat tolerance of Ganoderma lucidum by promoting mitochondrial respiration driven by fatty acid β-oxidation”。

背景与目的:
在全球气候变暖的背景下,高温已成为一个不可避免的环境因素,影响着生长、代谢和发育等多种生理过程。研究热应激的机制有助于阐明生物体在热胁迫下的代谢模式,并通过人工代谢干预提高其耐热性。在植物中,热胁迫的影响及其相关的耐受机制已有较为深入的研究。例如,在水稻开花期,热胁迫会严重降低结实率,而水稻可以通过脱落酸(ABA)信号通路抵抗热胁迫。长期热胁迫会引发番茄果实的程序性细胞死亡,而番茄细胞壁转化酶则能通过抑制与活性氧(ROS)无关的细胞死亡来增强耐热性。在微生物中,热胁迫深刻影响生长和代谢,而细胞则通过改变胞内代谢的来调节其在胁迫下适应能力。例如,热胁迫诱导酿酒酵母的细胞死亡,而酿酒酵母通过增强Atg32依赖的线粒体自噬增加其耐热性。在罗伯茨绿僵菌(Metarhizium robertsii)中,热胁迫显著抑制菌丝生长,而该真菌通过丙酮酸积累增强热应激耐受性。然而,关于大多数微生物的耐热机制的研究仍然有限。研究真菌的耐热机制有助于我们更好地理解微生物对热胁迫的代谢反应。
多胺是胁迫响应的关键调节因子,参与了多种生物过程的调控。在植物中,各种非生物胁迫,如高温、低温、干旱和盐胁迫,都会导致多胺的积累。亚精胺是多胺家族的重要成员,在多种胁迫耐受中起着重要作用。在植物中,亚精胺通过调节一系列细胞内代谢途径来增强热胁迫抗性。例如,亚精胺通过调节内源性淀粉和多胺代谢来提高水稻种子的耐热性,并通过调节γ-氨基丁酸(GABA)含量和代谢来增强白三叶草的耐热性。然而,关于亚精胺对真菌耐热性的影响及其潜在机制的研究却很少被报道。
线粒体在细胞稳态中起着至关重要的作用,并且是胁迫响应的关键组成部分。线粒体呼吸产生ATP,为防御机制提供能量,且其还能作为次级代谢的碳中间体来源。在拟南芥中,线粒体ATP合酶亚基D的敲低会导致ATP生成减少和耐热性下降。大量研究表明,多胺与线粒体功能密切相关。因此,生物体中可能存在一种潜在的调控途径,即亚精胺通过调节线粒体功能来增强生物体耐热性。
方法与结果:
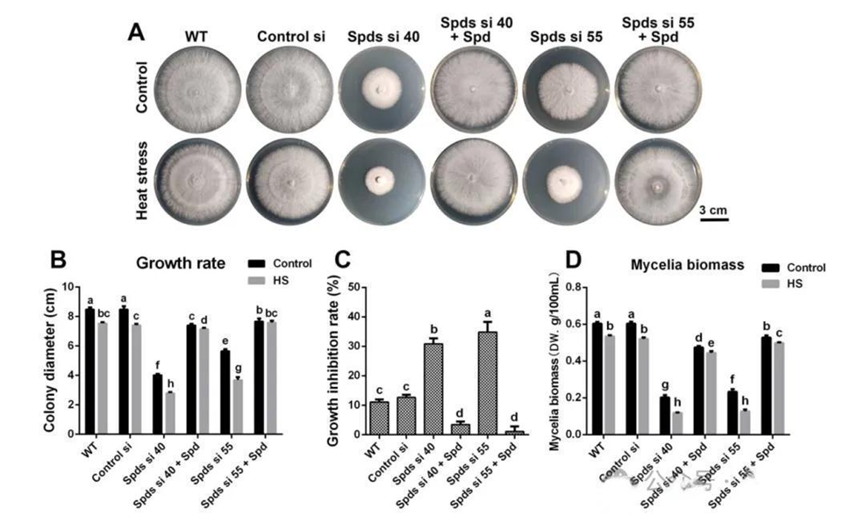
一、亚精胺增强灵芝的耐热性。亚精胺合成酶(SPDS)是参与亚精胺生物合成的关键酶。在敲低spdS后,灵芝中亚精胺的水平降低了60%以上。为了研究亚精胺对灵芝耐热性的影响,作者检测了野生型(WT)和spdS敲低菌株的生长速率。发现在42℃热胁迫处理30分钟后,所有基因型的生长速率均受到抑制。然而,野生型的生长抑制率约为11.1%,而spdS敲低菌株的生长抑制率超过30%,表明spdS敲低导致对热应激的敏感性增加。在spdS敲低菌株中,补充亚精胺后热应激诱导的抑制得到缓解。这些结果表明亚精胺显著增强了灵芝的耐热性。
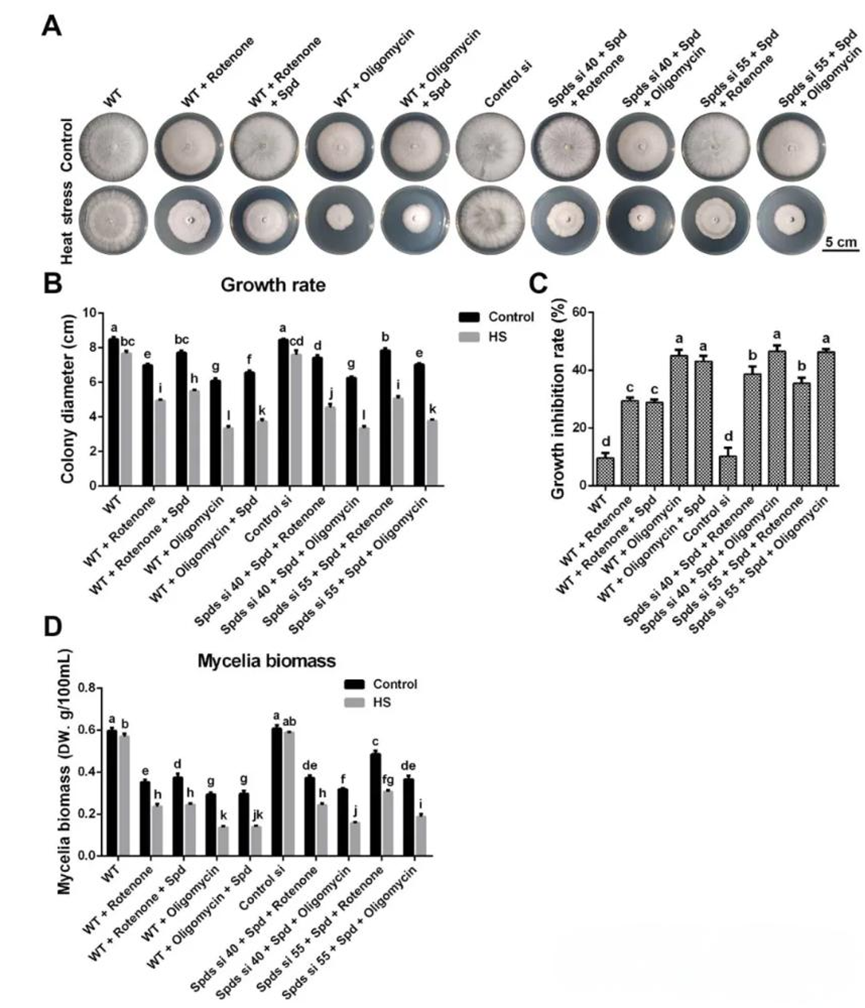
二、线粒体呼吸对亚精胺增强灵芝耐热性至关重要。线粒体在细胞稳态中起着至关重要的作用,并且是胁迫响应的关键组成部分。为了确定亚精胺是否通过调控线粒体功能增强灵芝的耐热性,作者首先检测了线粒体呼吸作用。结果显示,野生型的OCR在热应激30分钟后增加了近80%,但在敲低spdS基因后,这种增加被抑制。在spdS敲低菌株中,OCR比野生型降低了70%-75%,而在补充1 mM亚精胺后,OCR恢复到野生型水平,ATP含量也表现出与OCR相似的趋势。这些结果表明,亚精胺促进了热胁迫诱导的线粒体呼吸。随后,通过使用鱼藤酮(线粒体复合物I抑制剂)和寡霉素(ATP合酶抑制剂)抑制线粒体呼吸,发现野生型对热胁迫的敏感性在鱼藤酮或寡霉素处理后显著增加,而补充亚精胺后耐热性并未显著改善。在spdS敲低菌株中,补充亚精胺后热胁迫诱导的抑制得到缓解,但在鱼藤酮或寡霉素处理后未观察到这种效果。这些结果表明线粒体呼吸对于亚精胺增强灵芝的耐热性至关重要。
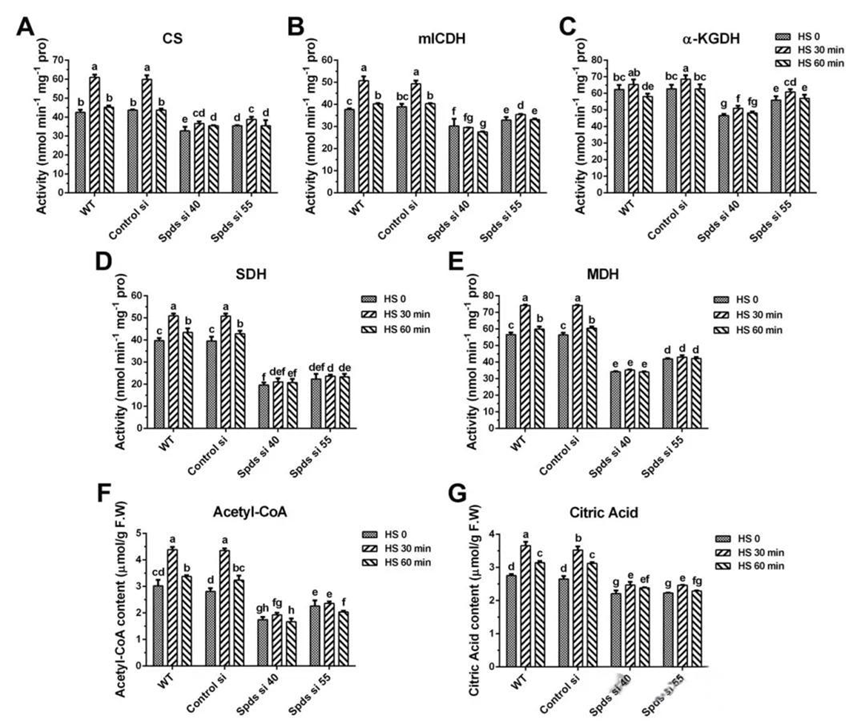
三、亚精胺通过加速TCA循环和促进电子传递来增强线粒体呼吸。为了探索线粒体对热胁迫的响应以及亚精胺在线粒体呼吸中的调控作用,作者分析了线粒体复合物的活性以及线粒体NAD/NADH和细胞内FAD/FADH2的比值。结果显示在野生型中,热胁迫30分钟后,复合物I、II、III和IV的活性显著增加。然而,在敲低spdS基因后,复合物I和II的活性受到抑制,且热胁迫诱导的增加消失。对野生型中线粒体NAD/NADH和细胞内FAD/FADH2的分析显示,在热应激30分钟后,NAD/NADH和FAD/FADH2的比值分别下降了41.2%和33.5%,表明热应激促进了NADH和FADH2的积累。与野生型相比,spdS敲低菌株的NAD/NADH和FAD/FADH2比值分别增加了2.1倍和1.7~2.0倍,表明spdS敲低降低了NADH和FADH2的水平。然而,这一结果与spdS敲低会通过抑制复合物I和II诱导NADH和FADH2积累的假设不一致。因此,spdS敲低可能也抑制了NADH和FADH2的产生。线粒体NADH由TCA循环产生,而FADH2由TCA循环和脂肪酸β-氧化产生。随后,作者分析了TCA循环中关键酶的活性和关键底物的水平。在热胁迫30分钟后,野生型中柠檬酸合酶(CS)、线粒体异柠檬酸脱氢酶(mICDH)、苹果酸脱氢酶(MDH)和琥珀酸脱氢酶(SDH)的活性分别增加了43.5%、34.4%、31.0%和28.3%。在spdS敲低菌株中,CS、mICDH、MDH和SDH的活性与野生型相比显著降低,且热胁迫后这些酶的活性几乎没有显著增加。此外,在热胁迫下,野生型中α-酮戊二酸脱氢酶(α-KGDH)的活性没有显著变化,但在spdS敲低菌株中却受到抑制。对关键底物水平的分析显示,在野生型中,热胁迫30分钟后乙酰辅酶A(Ac-CoA)和柠檬酸(CA)的水平增加了30%~40%,但在spdS敲低菌株中,热胁迫下它们的含量分别下降了46.2%~56.1%和32.5%。综上所述,这些结果表明线粒体响应热胁迫,而亚精胺通过加速TCA循环和促进电子传递来增强热胁迫下的线粒体呼吸。
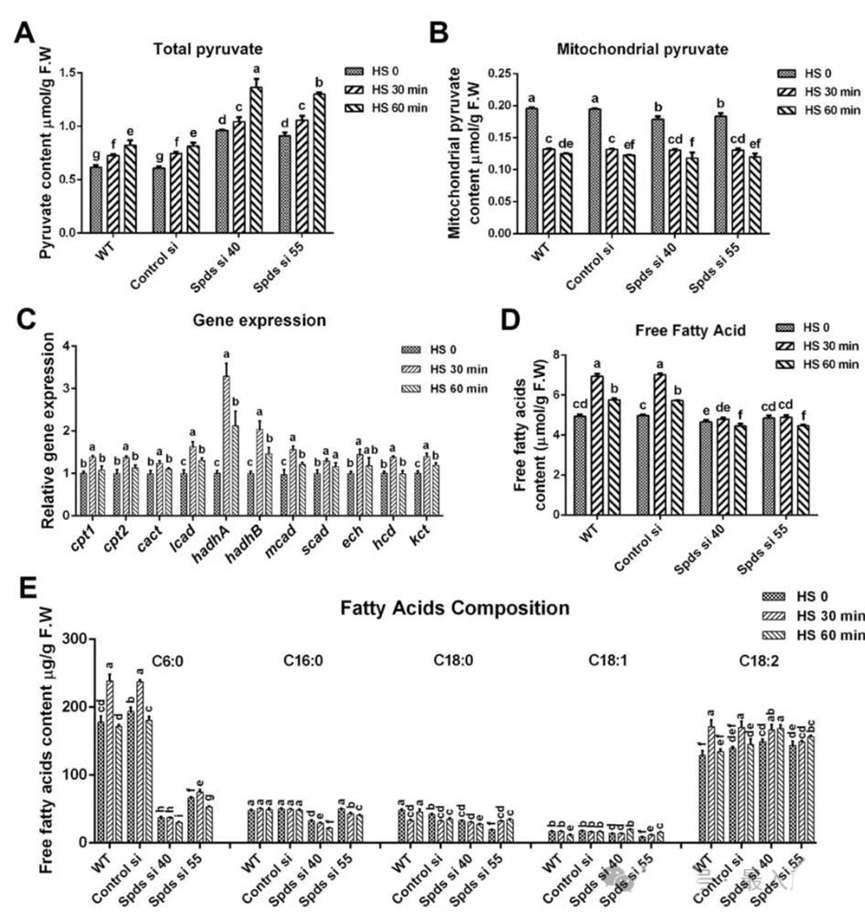
四、亚精胺增强热胁迫诱导的脂肪酸β-氧化。TCA循环产生电子载体NADH和FADH₂,随后被电子传递链(ETC)利用。由丙酮酸转化形成的乙酰辅酶A(Ac-CoA)是启动TCA循环反应的主要底物。为了确定热胁迫下线粒体呼吸的增强是否由丙酮酸驱动,作者检测了总丙酮酸和线粒体丙酮酸的水平。在野生型中,总丙酮酸在热胁迫下逐渐积累,热胁迫60分钟后增加了33.6%。相反,野生型中线粒体丙酮酸水平在热胁迫60分钟后下降了36.1%。此外,在热胁迫下,spdS敲低菌株中的总丙酮酸水平比野生型增加了56.8%~66.0%,但线粒体丙酮酸水平与野生型相当。这些结果表明,热胁迫下丙酮酸进入线粒体受阻,而spdS敲低促进了丙酮酸的积累,但对丙酮酸进入线粒体的影响较小。
由脂肪酸β-氧化产生的乙酰辅酶A在有氧呼吸过程中直接进入TCA循环,而脂肪酸β-氧化是FADH₂产生的主要来源。为了确定热胁迫诱导的线粒体呼吸是否由脂肪酸β-氧化介导,作者评估了与脂肪酸β-氧化相关基因的表达水平。在热应激下,野生型中脂肪酸β-氧化相关基因的表达水平显著增加。此外,对游离脂肪酸含量的分析表明,野生型在热应激30分钟后游离脂肪酸水平增加了28.8%,但在spdS敲低菌株中未观察到这种增加,其水平显著低于野生型。随后,分析了游离脂肪酸的组成。在野生型中,热胁迫30分钟后中链脂肪酸(C6:0)的含量增加了33.9%,而在spdS敲低菌株中,热胁迫下其含量下降了68.6%~84.2%。然而,长链脂肪酸(C18:2)的含量在spdS敲低菌株中略有升高。综上所述,这些发现表明亚精胺增强了热胁迫诱导的脂肪酸β-氧化。
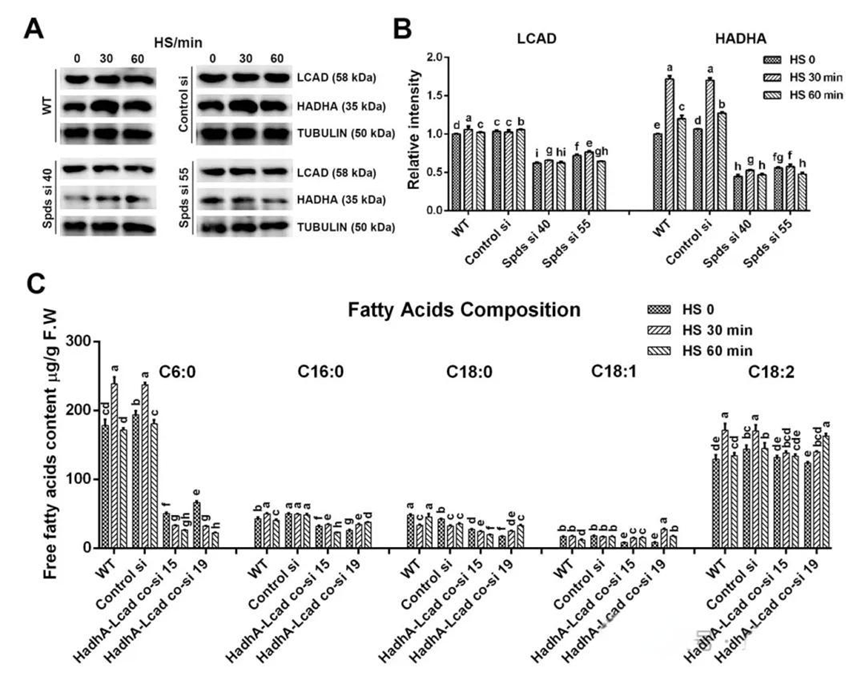
五、亚精胺促进LCAD和HADH的翻译。长链脂肪酸β-氧化的抑制(例如CPT、LCAD或HADH的缺陷)会导致长链脂肪酸的积累和短/中链脂肪酸的减少。为了探索亚精胺在热胁迫下对长链脂肪酸β-氧化的影响,作者评估了与长链脂肪酸转运和β-氧化相关的基因转录水平及多聚核糖体mRNA比例。结果表明,在热应激下,野生型中与长链脂肪酸转运和β-氧化相关基因的转录水平增加,特别是lcad、hadhA和hadhB,但在敲低spdS后未受到显著影响。然而,lcad、hadhA和hadhB的多聚核糖体mRNA比例在敲低spdS基因后受到显著抑制。随后,作者分析了HADH和LCAD的蛋白水平,野生型在热胁迫30分钟后HADH增加了72%。然而,敲低spdS后热胁迫诱导的增加被抑制,其蛋白水平显著低于野生型。在热胁迫下,野生型中LCAD水平几乎没有增加,但在spdS敲低菌株中显著下降。这些结果表明,亚精胺主要通过增加lcad、hadhA和hadhB的多聚核糖体mRNA比例,来促进热胁迫下LCAD和HADH的翻译。随后,作者构建了lacd和hadhA基因共敲低菌株,对游离脂肪酸组成的分析显示,lacd和hadhA共敲低菌株中中链脂肪酸(C6:0)水平显著降低,而长链脂肪酸(C18:2)的比例增加,这与spdS敲低菌株的趋势相似。综上所述,亚精胺通过促进热胁迫下LCAD和HADH的翻译来加速脂肪酸β-氧化。
结论:
多胺是存在于所有生物体中的重要有机阳离子,是胁迫响应的关键调节因子,参与了多种生物过程的调控。在本研究中,线粒体呼吸在亚精胺增强灵芝耐热性中起着关键作用。分析表明,亚精胺增强了TCA循环和ETC关键酶的活性,并通过促进LCAD和HADH的翻译参与热胁迫诱导的脂肪酸β-氧化。基于这些结果,作者提出了亚精胺通过促进线粒体呼吸增强耐热性的潜在细胞级联反应。此研究揭示了亚精胺增强灵芝耐热性的初步机制,为真菌中多胺在非生物胁迫下的作用提供了新的见解。
 上一篇:László G. Nagy团队在Current biology杂志发表大型真菌孢子形成的新见解
上一篇:László G. Nagy团队在Current biology杂志发表大型真菌孢子形成的新见解- 下一篇:Mycology | 黄晨阳团队揭示PoMbp1转录因子通过调控多糖利用促进糙皮侧耳的生长发育

