
Cell Discovery|酵母中蘑菇衍生生物活性II型灵芝酸生物合成网络的解码和重编程


摘要
蘑菇来源的次级代谢产物具有重要的药理活性,但其生物合成途径的阐明极具挑战性,更不用说将其生物合成网络重新编程为目标代谢物。本文以传统药用真菌灵芝(Ganoderma lucidum)为例,通过研究灵芝中基因的协同表达,揭示了灵芝主要活性代谢产物II型灵芝酸(TIIGAs)的生物合成网络,鉴定了能够负责C22羟基化、C3羟基构型转化和C3、C15和C22羟基上的乙酰化的内源或异源酶。值得注意的是,我们揭示了C22羟化酶CYP512W6的催化机制,以及将乙酰基转移到C15和C22所需的意想不到的双功能乙酰转移酶GlAT。使用荧光引导的整合方法,我们实现了适用于工业发酵的重要TIIGA的高效生物合成。在将所有鉴定的酶引入工程酵母后,我们观察到下游TIIGA的生物合成受到严重阻碍,并且通过暂时调节乙酰转移酶的表达来疏通代谢阻断。通过重新编程TIIGA的生物合成网络,我们能够生产超过30种TIIGA,其滴度或效率比灵芝栽培高出1-4个数量级。这项工作使人们能够获得有价值的TIIGA,促进其广泛应用,并为其他蘑菇产品的研究提供了启示。
杂志名:Cell Discovery
英文题目:Decoding and reprogramming of the biosynthetic networks of mushroom-derived bioactive type II ganoderic acids in yeast
作者:Qin Wang,YeLi, Shun han Zhang, Wei Yuan , Ze qian Du, Ting Shi, Zhao Chang, Xing ye Zhai, Yin hua Lu, Meng Wang, Juan Guo, Jian-Jiang Zhong and Han Xiao
通讯作者单位:上海交通大学
内容
1. 缩小后修饰基因的候选范围
使用灵芝菌丝体粗蛋白裂解物,以GA-Jb作为底物进行体外活性测定。液相色谱-质谱(LC-MS)分析显示,生成了化合物1,表明,C3羟基的构型转化可以通过两步反应来完成,即C3羟基的氧化,然后C3酮基的还原。采用蓝光暴露策略来特异性刺激菌丝体中TIIGA的产生,随后进行RNA测序。使用CYP5150L8和CYP512W2作为共表达分析的诱饵基因,发现168个转录物与它们的表达高度相关。基于与TIIGA的底物结构相似性,最终选择了12种CYP、8种AT和11种脱氢酶/还原酶。
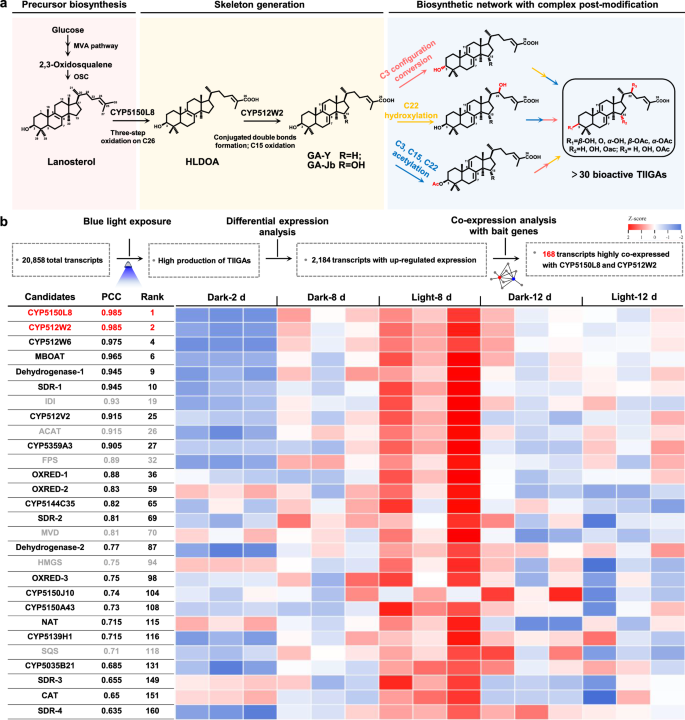
图1 灵芝中TIIGAs的生物合成网络分析
2. C3羟基构型转化的异源氧化酶和还原酶的鉴定
为了鉴定负责C3羟基构型转化的酶,将异源脱氢酶候选物引入到GA-Jb生产菌,烟曲霉(AfuSDR)和甜橙(CsSDR)的短链脱氢酶/还原酶能够催化GA-Jb的转化以产生GA-TR(图2a)。来自智人的AKR1C2和AKR1C4以及来自纺锤枝顶孢霉的FusC1能够将GA-TR完全转化为GA-Ja(图2b)。将两种氧化酶(CsSDR和AfuSDR)和三种还原酶(AKR1C2、AKR1C4和FusC1)随机分组为六种氧化酶-还原酶对,并将它们与CYP512W2一起引入HLDOA生产酵母中。CsSDR和AKR1C4的共表达产生33.3mg/L GA-Ja并积累5.4mg/L GA-Jb,没有可检测的GA-TR(图2c)。C3羟基的氧化还原介导的构型转化可以应用于其他GA,但其对HLDOA和GA-Y的C3羟基的转化不如GA-Jb有效。
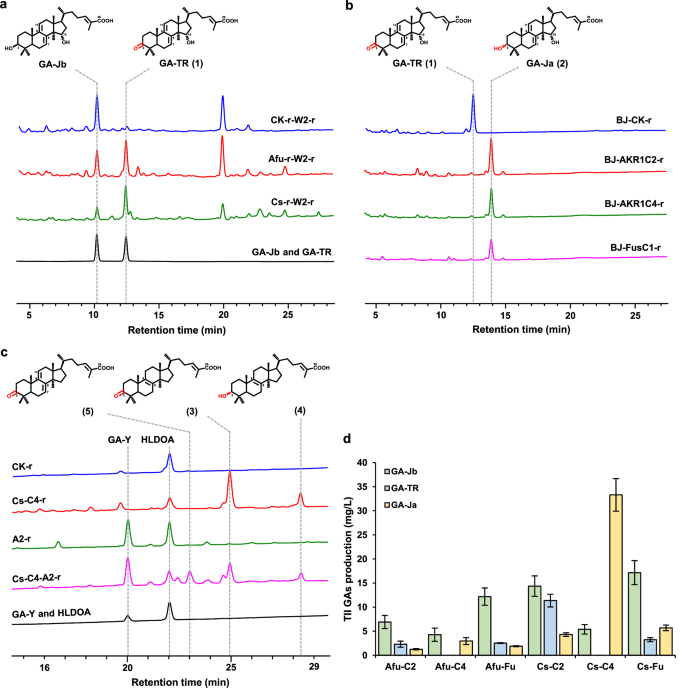
图2 C3氧化酶和还原酶的功能特性
3. CYP512W6作为C22羟化酶的鉴定及其催化机制的阐明
体内异源表达发现CYP512W6可以将HLDOA、GA-Y和GA-Jb的C22位羟基化。酵母微粒体体外反应验证CYP512W6只能催化GA-Jb上的C22羟基化,而不能催化其他GA上的C22羟基化,GA-Jb是CYP512W6的最佳底物(图3a,b,c,d)。为了研究CYP512W6介导的GA-Jb上C22羟基化的催化机制,通过AlphaFold 2对CYP512W6的结构进行建模。发现除了催化口袋周围残基的变化外,CYP512W6和CYP512W2的总体结构相似。量子力学(QM)计算和分子动力学(MD)模拟共同证明GA-Jb可以被CYP512W6羟基化(图3e)。基于MD模拟,我们鉴定了8个残基(V105、L108、L109、T212、S363、L364、M365、R366),结构分析表明,T212的侧链和M365的主链分别与GA-Jb的C3羟基和C26羧基形成氢键,而V105、L108和M365则与GA-Jb的四环发生疏水作用(图3g)。此外,S363与GA-Jb的C15羟基形成硫-氢键,R366与血红素的羧基形成盐桥。设计五个突变体(V105 L,L108 A,T212 A,M365 A和R366 A),发现所有突变体都表现出对GA-Jb的活性显著降低(图3h)。
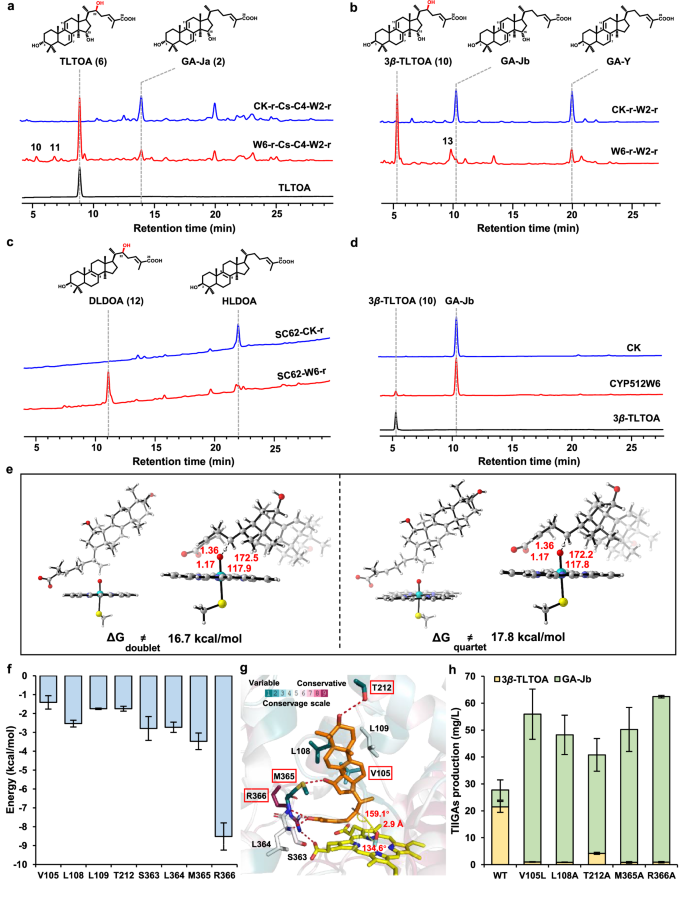
图3 CYP512W6作为C22羟化酶的发现
4.GlAT作为C15和C22羟基上双功能乙酰转移酶的鉴定及其催化机制的阐明
体内异源表达GlAT,以GA-Jb和GA-Ja为底物,检测到C15乙酰化的产物。制备GlAT的酵母微粒体,并将它们与不同的底物一起孵育。发现它可以催化各种底物的C15羟基的乙酰化。表明GlAT是能够在C15和C22上乙酰化的双功能乙酰转移酶。使用AF2对GlAT进行结构预测。对接结果表明,GA-Jb的O 1与酰基辅酶A的Cα之间的距离以及H305的Nε与GA-Jb的O 1之间的距离均为3.1 nm,适合乙酰化(图5 c)。MD模拟结果证明了GlAT在C15和C22位置转移酰基的能力。根据MD模拟结果,观察到T型隧道周围的G208和A211为GA-T2入口提供空间。S79和R218与GA-T2尾部的相互作用影响乙酰化位置(图5 b)。因此,S79 L、G208 L、A211 L、A211 F和R218 L被设计为改变T-通道大小或破坏相互作用以改变酰化位点。
H305 A在维持C15和C22上的乙酰化活性方面的关键作用。A211可能是双功能乙酰化过程的另一个重要残基,G208可能负责维持C22乙酰化活性。一种可能的解释是,208位的亮氨酸比同一位置的甘氨酸产生更大的空间位阻。这种空间位阻的差异不同地影响具有不同结构特征的底物,导致不同底物在相同化学位置处的不同选择性。推断双官能乙酰化活性可能受到C3、C15和/或C22上的全面修饰的影响。A211 F突变可能完全阻断了Ttunnel,阻止底物进入催化位点,而A211 L突变虽然也减少了T-tunnel空间,但在接近催化位点时仍为GA-T2的C22乙酰化留下了足够的空间。然而,剩余的空间不足以使C15乙酰化所需的进一步向内移动。
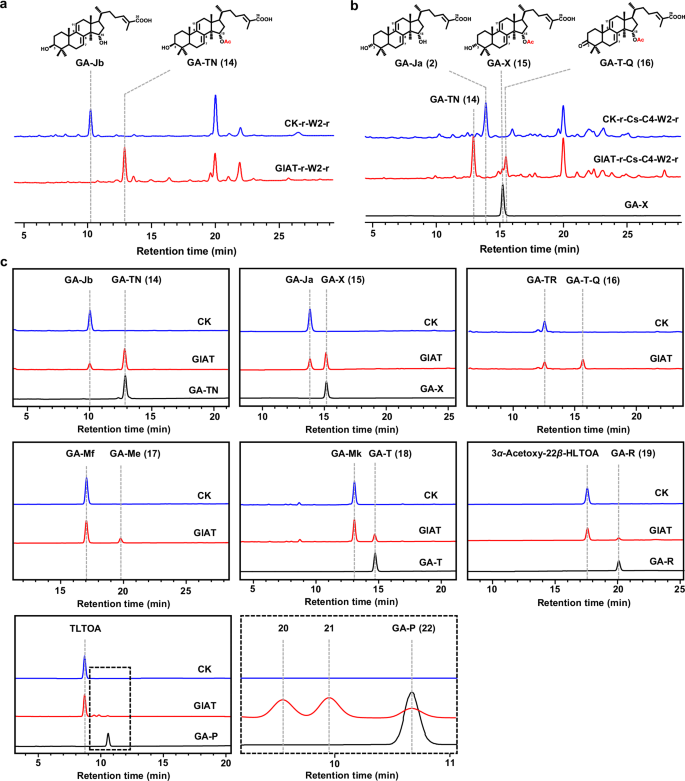
图4 GlAT的功能表征
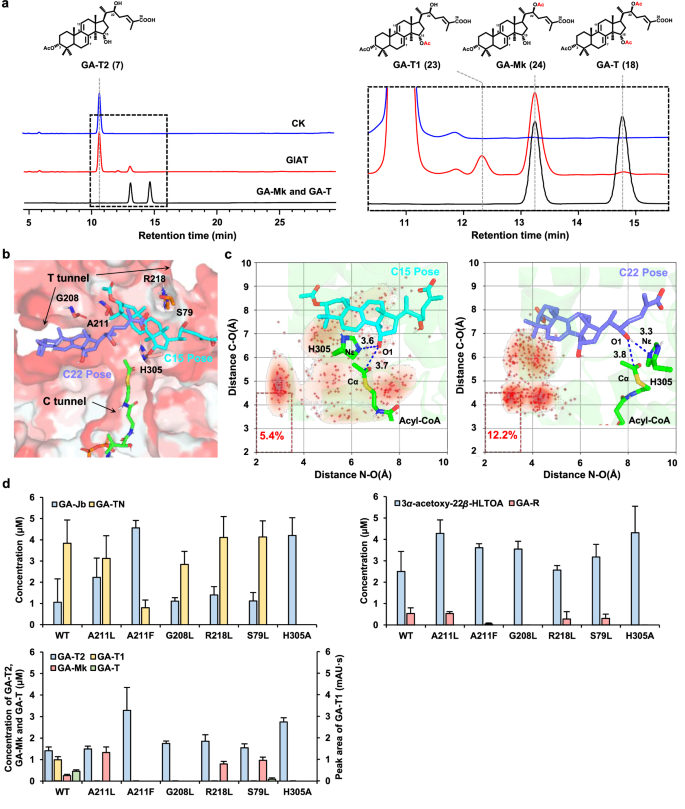
图5发现GlAT作为C15和C22上的双功能乙酰转移酶
5.异源BsAT作为C3羟基乙酰转移酶的鉴定
异源表达证明BsAT能够乙酰化TIIGA的不同C3羟基构型。BsAT对其他TIIGA(DLTOA、GA-S、TLTOA、GA-P)具有催化活性,而不管后修饰(羟基化或乙酰化)是否发生在C15和/或C22位置。BsAT具有底物偏好性,GA-S是最有利的,能够完全转化。与C3β-羟基底物相比,BsAT对C3α-羟基底物具有较高的催化活性,对C15-乙酰化TIIGA和C22-羟基化TIIGA具有较高的转化效率。
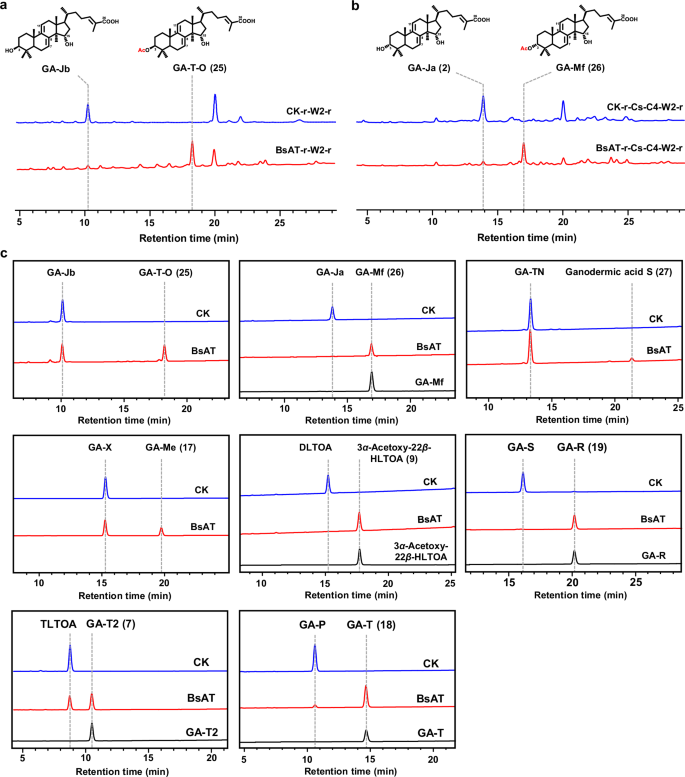
图6 BsAT的功能表征
6. 重编程TIIGAs的生物合成网络
通过荧光引导整合技术将催化模块整合进酵母基因组。并通过荧光激活细胞分选(FACS)进行筛选(图7a)。在不添加抗生素的情况下,菌株SC27能够产生最高量的GA-Jb,为39.6 mg/L(图7 b)。将其进一步在含有1g/L 5-FOA的YPD培养基中进行培养以回收CRISPR质粒。最终得到的菌株SC27*能够在120小时发酵后产生41.1 mg/L的GA-Jb(图8)。
将GA-Jb生产菌株SC27*工程化以生产GA-Ja和TLTOA。菌株SC 3和SC 1能够在120小时发酵后分别产生最高量的GA-Ja(87.5mg/L)和TLTOA(37.4mg/L),而不添加抗生素。经质粒回收后,最终得到的菌株SC 3 * 和SC 1 * 在发酵120 h后分别能够产生47.0 mg/L的GA-Ja和29.2 mg/L的TLTOA。
通过使用诱导型启动子来驱动BsAT和/或GlAT的表达来控制后修饰反应的顺序。选择半乳糖诱导型启动子PGAL1来驱动BsAT基因表达,促进C3位的乙酰化反应。采用了PADH2(乙醇诱导型启动子)和PGAL1来控制GlAT的表达,延缓C15与C22位乙酰化反应,以积累中间产物GA-T。最终通过工程酵母菌株的改造,超过30种TIIGA的生产效率或产量比来自养殖蘑菇的生产效率或产量高1-4个数量级。
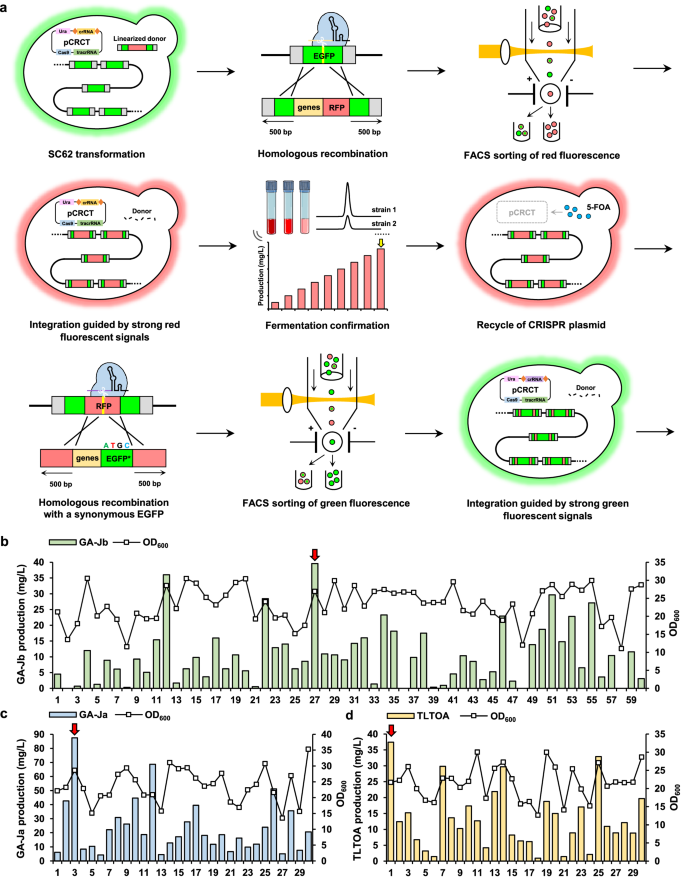
图7关键酶表达盒的整合导致TIIGA的有效产生
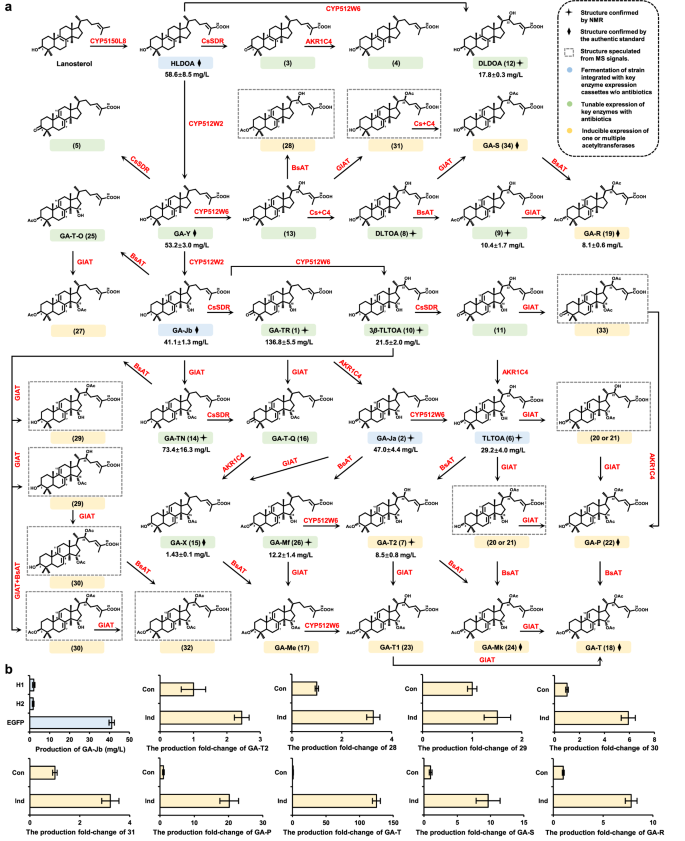
图8将生物合成网络重编程为靶向TIIGA

